The gnathostomes, or jawed vertebrates (Infraphylum Gnathostomata, Subphylum Vertebrata) comprise all living vertebrates except for the Cyclostomata.
The stem group of the Gnathostomes is very well represented in the fossil record. Many phylogenetic studies have been carried out, one of the most recent and comprehensive of which is that by Sallan et al (2018). The following phylogenetic time tree (split into two parts) is based, with the modification mentioned in the figure, on the tree in their article:
The stem group of the Gnathostomes is very well represented in the fossil record. Many phylogenetic studies have been carried out, one of the most recent and comprehensive of which is that by Sallan et al (2018). The following phylogenetic time tree (split into two parts) is based, with the modification mentioned in the figure, on the tree in their article:
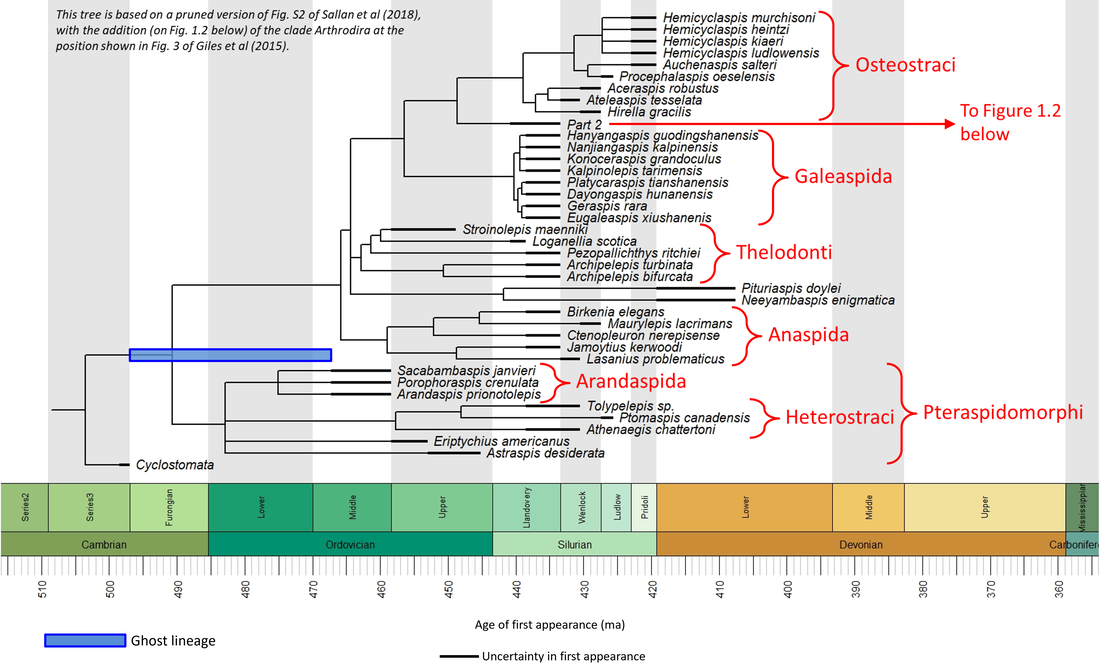
Figure 1.1. Time tree of the stem-Gnathostomata (Part1)
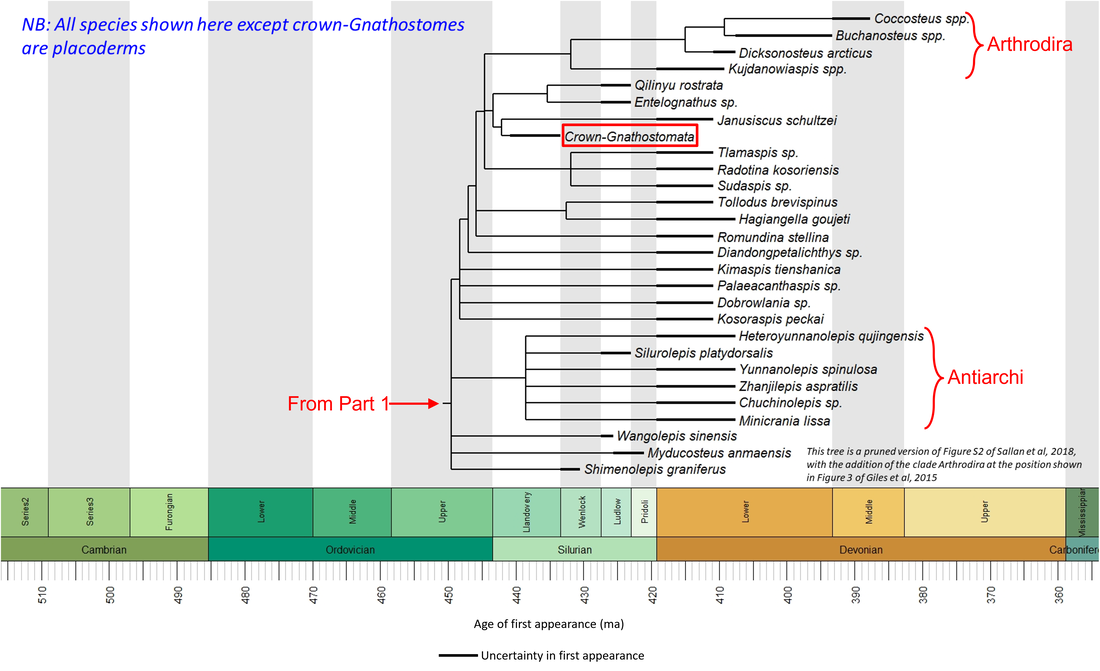
Figure 1.2. Time tree of the stem-Gnathostomata (Part2)
The above tree represents only a small proportion of the total number of stem-group gnathostomes. Many lie far out on the side branches of the above tree, phylogenetically distant from the main stem line leading to the crown group; these are less informative as transitional fossils than those that lie close to the main stem line, and are thus excluded from consideration here.
The stem-Gnathostomata appeared during the Darriwilian age of the Middle Ordovician. Three genera are known from that time interval: Arandaspis, Sacabambaspis and Pteraspis. Of these, the first two have been described at the species level, and can be considered as the oldest-known representatives of the Stem-Gnathostomata:
Both species are illustrated in Figure 2 below..
Owing to the large number of stem-Gnathostomata, we will deal with the stem line in the following stages (refer to the clade names shown in the phylogenetic tree above):
Only the species crownward of the Osteostraci have jaws. These belong to the polyphyletic group known as placoderms.
Pteraspidomorphi
Examples of species from this most basal part of the phylogenetic tree are shown below (click on image for a larger view; this applies to all fossil image sets further below on this page):
The stem-Gnathostomata appeared during the Darriwilian age of the Middle Ordovician. Three genera are known from that time interval: Arandaspis, Sacabambaspis and Pteraspis. Of these, the first two have been described at the species level, and can be considered as the oldest-known representatives of the Stem-Gnathostomata:
- Arandaspis prionotolepis, a primitive jawless fish, belonging to the Arandaspida (the sister group of the Heterostraci), from the Middle Ordovician Stairway Sandstone of Mount Watt and Mount Charlotte, Northern Territory, Australia (Ritchie and Gilbert-Tomlinson, 1977; Benton et al, 2015).
- Sacabambaspis janvieri, another jawless fish belonging to the Arandaspida and found in the Middle Ordovician Anzaldo Formation at Sacabamba in the department of Cochabamba, Bolivia (Gagnier et al, 1986; Sallan et al, 2018).
Both species are illustrated in Figure 2 below..
Owing to the large number of stem-Gnathostomata, we will deal with the stem line in the following stages (refer to the clade names shown in the phylogenetic tree above):
- Pteraspidomorphi
- Anaspida
- Thelodonti and their sister species
- Galeaspida
- Osteostraci
- Antiarchi and their sister species
- The remaining, most crownward, stem-group species
Only the species crownward of the Osteostraci have jaws. These belong to the polyphyletic group known as placoderms.
Pteraspidomorphi
Examples of species from this most basal part of the phylogenetic tree are shown below (click on image for a larger view; this applies to all fossil image sets further below on this page):



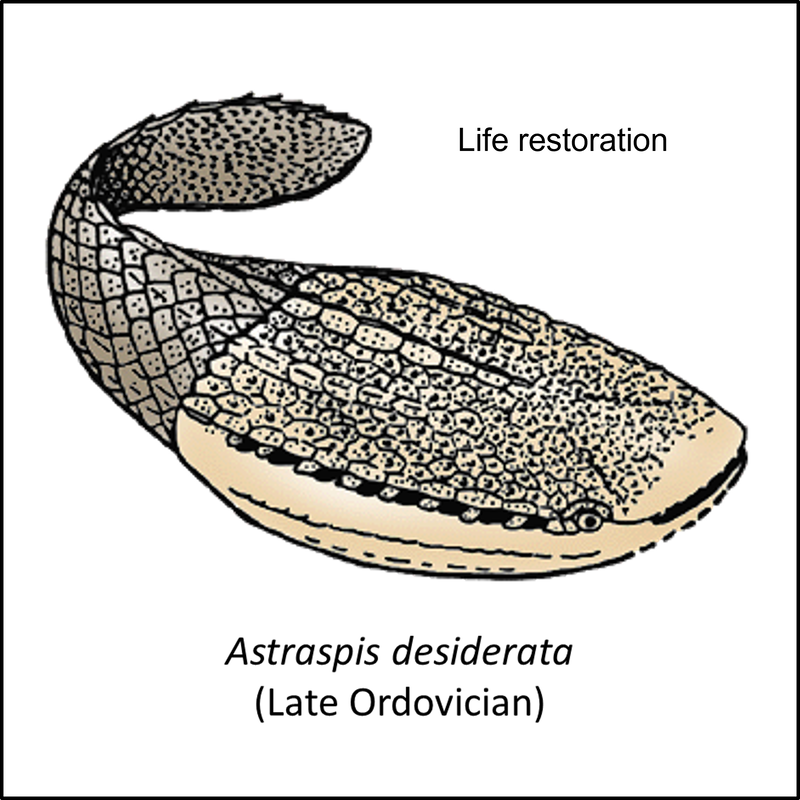


Figure 2. Images of stem-Gnathostomata: Pteraspidomorphi
The above images, together with those in the following image sets for the gnathostomes, are placed in left-to-right order from the most basal to the furthest crownward, according to the analysis of Sallan et al (2018). No obvious trends can be seen, except possibly for the hint of a progressively greater head width relative to the width of the rest of the body.
Anaspida
Some examples of the anaspid stem-Gnathostomata are illustrated in the following images:
Anaspida
Some examples of the anaspid stem-Gnathostomata are illustrated in the following images:




Names in red indicate that the fossil is younger than the oldest known crown-group fossil.
Figure 3. Images of stem-Gnathostomata: Anaspida
These species are also jawless and armored, but they have a form that more closely resembles that of modern fish. One characteristic of the anaspids, lacking in the pteraspidomorphs, that makes them bear a superficial resemblance to modern fishes is the fact that they possess paired fins (Janvier, 1987). Most do not have the massive head-shield seen in many of the more basal forms.
Note that one of the fossils illustrated above (Lasanius problematicus), and many in the illustrations below, have red labels. This indicates that their first appearance age is less than that of the gnathostome crown group, which appeared in the Early Silurian. These post-crown stem-group fossils represent descendants of ancestors that would have separated from the stem line during or before the Early Silurian.
Thelodonti and their sister species
These stem-group gnathostomes comprise the clade Thelodonti and its sister species. As with the anaspids, the armor of the thelodonts comprises scales, but these were generally dispersed after death, such that their anatomy is poorly understood (Donoghue and Smith, 2001). The images available in the public domain are shown below in Figure 4:
Note that one of the fossils illustrated above (Lasanius problematicus), and many in the illustrations below, have red labels. This indicates that their first appearance age is less than that of the gnathostome crown group, which appeared in the Early Silurian. These post-crown stem-group fossils represent descendants of ancestors that would have separated from the stem line during or before the Early Silurian.
Thelodonti and their sister species
These stem-group gnathostomes comprise the clade Thelodonti and its sister species. As with the anaspids, the armor of the thelodonts comprises scales, but these were generally dispersed after death, such that their anatomy is poorly understood (Donoghue and Smith, 2001). The images available in the public domain are shown below in Figure 4:





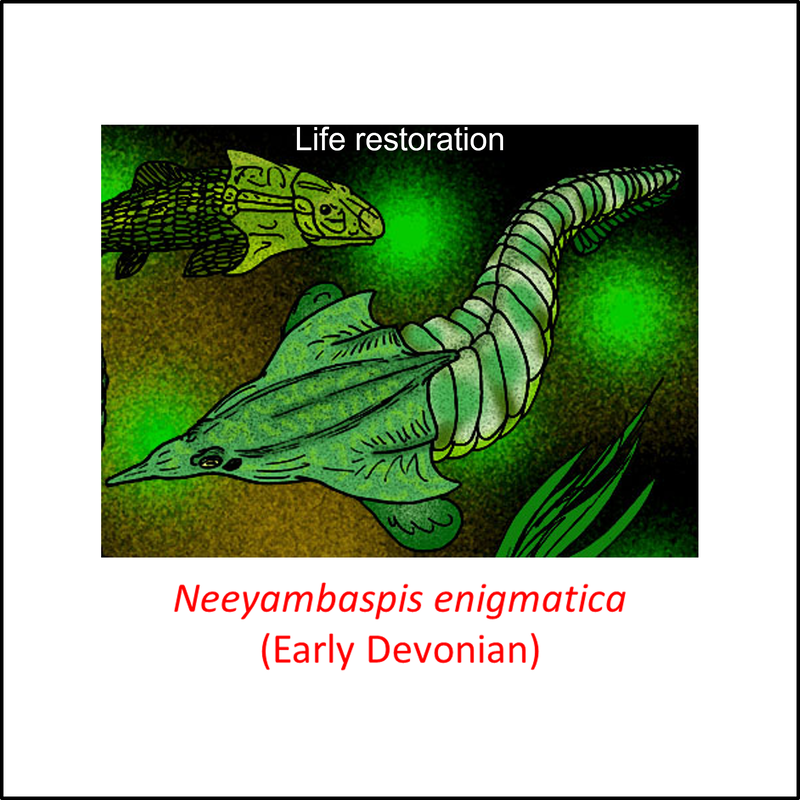
Names in red indicate that the fossil is younger than the oldest known crown-group fossil.
Figure 4. Images of stem-Gnathostomata: Thelodonti and sister species
These species are, like the more basal forms, jawless and armored, but they do not show any clear trend through the series shown above. Note the different head shape in the two species that are in a sister relationship to thelodonts (Pituriaspis and Neeyambaspis).
Galeaspida
This jawless clade, illustrated by the following images, comprises species that all have massive head-shields:
Galeaspida
This jawless clade, illustrated by the following images, comprises species that all have massive head-shields:


Figure 5. Images of stem-Gnathostomata: Galeaspida
The shape and nature of the head-shield is superficially similar to the shield of many of the Heterostraci (for example, Athenaegis and Tolypelepis in Figure 2).
Osteostraci
The next clade to separate from the gnathostome stem line was the Osteostraci, another group of jawless fish. They are generally similar to the galeaspids, but their bone type is cellular (Janvier, 1984) while at least part of the bone in the galeaspids is acellular (Keating et al, 2018) and the bone structure of the head differs in detail. Here are some examples:
Osteostraci
The next clade to separate from the gnathostome stem line was the Osteostraci, another group of jawless fish. They are generally similar to the galeaspids, but their bone type is cellular (Janvier, 1984) while at least part of the bone in the galeaspids is acellular (Keating et al, 2018) and the bone structure of the head differs in detail. Here are some examples:

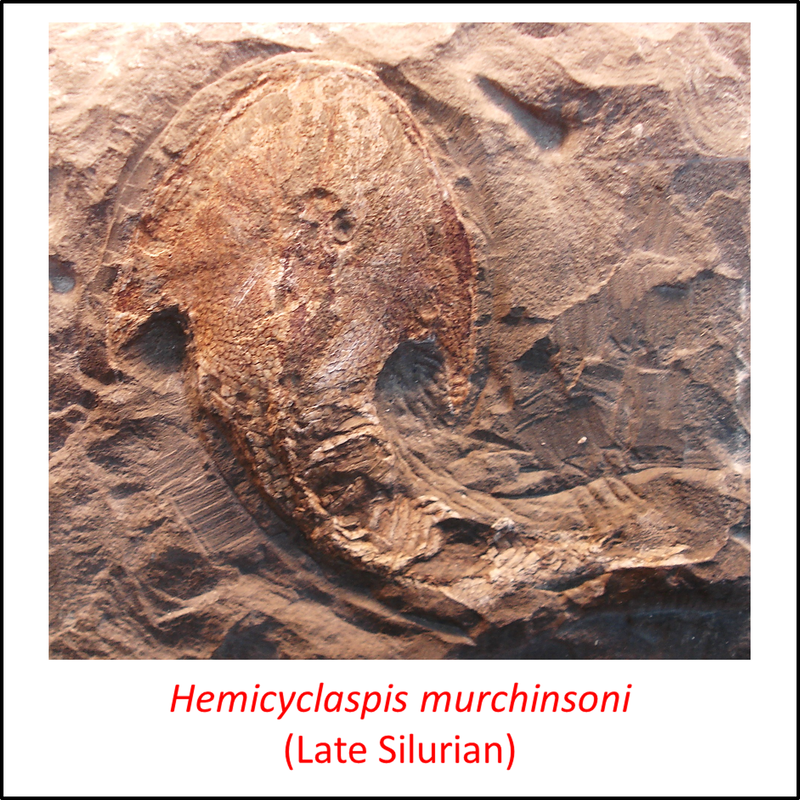

Names in red indicate that the fossil is younger than the oldest known crown-group fossil.
Figure 6. Images of stem-Gnathostomata: Osteostraci
The more basal species shown above have a head-shield with a characteristic pointed lateral extension on each side .
Antiarchi and their sister species
This group together with all the more crownward species comprise the placoderms (the jawed stem gnathostomes; Zhu et al, 2016). Species for which public-domain images are available are illustrated below:
Antiarchi and their sister species
This group together with all the more crownward species comprise the placoderms (the jawed stem gnathostomes; Zhu et al, 2016). Species for which public-domain images are available are illustrated below:




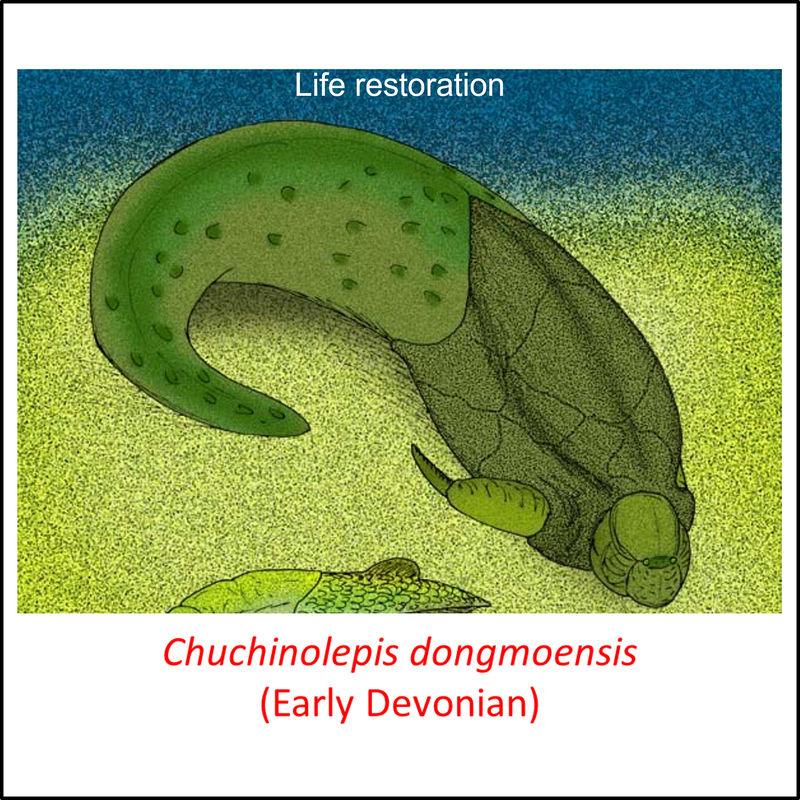

Names in red indicate that the fossil is younger than the oldest known crown-group fossil.
Figure 7. Images of stem-Gnathostomata: Antiarchi and sister species
Besides having jaws, the antiarchs and their sister species are also different from more basal stem gnathostomes in being more heavily armored, with a box-like thoracic shield and elongate pectoral fins encased in bone (Zhu et al, 2012). A good example of the bony pectoral fins can be seen above in Yunnanolepis chii.
The remaining, most crownward, stem-group species
These species represent placoderms that progressively separated from the gnathostome stem line crownward of the antiarchs. Some examples are shown below:
The remaining, most crownward, stem-group species
These species represent placoderms that progressively separated from the gnathostome stem line crownward of the antiarchs. Some examples are shown below:

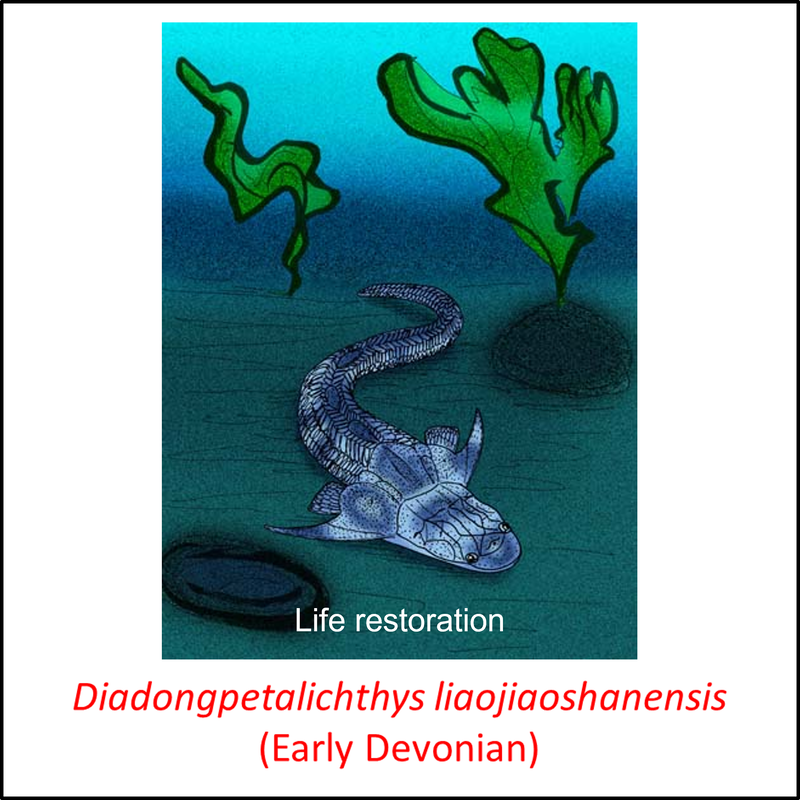
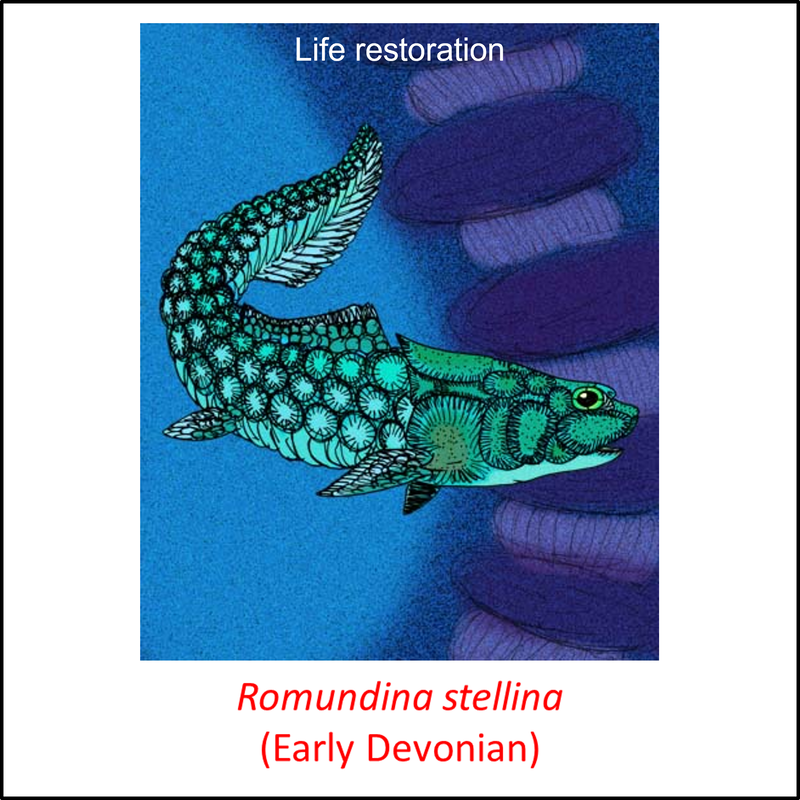


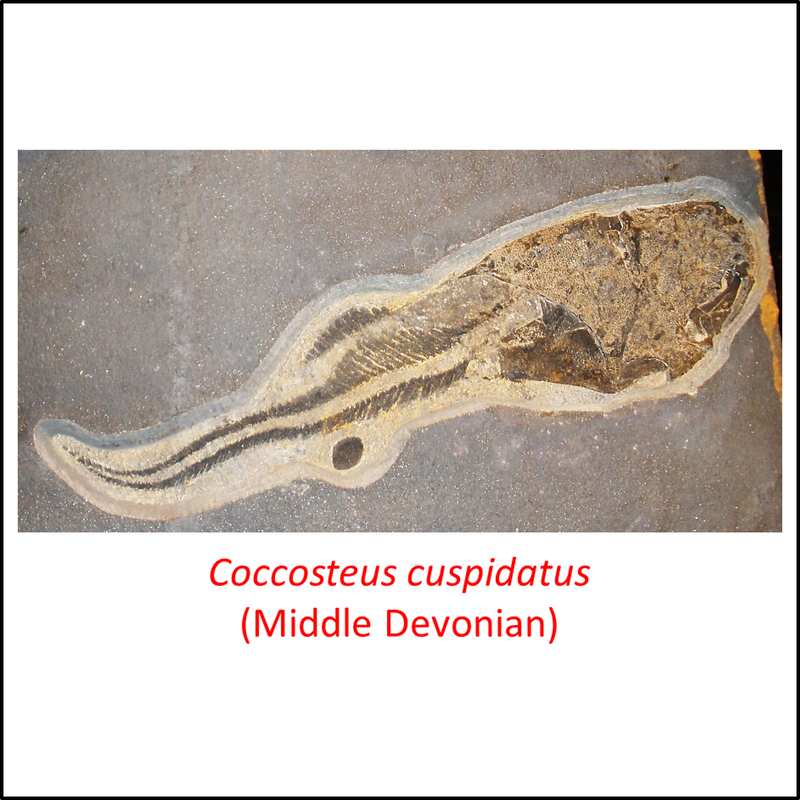




Names in red indicate that the fossil is younger than the oldest known crown-group fossil.
Figure 8. Images of stem-Gnathostomata: The remaining, most crownward, stem-group species
These species are characterized by armor that comprises large plates and scales. Four of the fossils shown above (Kujdanowiaspis, Dicksonosteus arcticus, Coccosteus cuspidatus and Buchanosteus confertituberculatus) belong to the Arthrodira. This clade differs from more basal stem-Gnathostomata in having a movable joint between the head and the body armor. Like the antiarchs, they also had teeth (Rücklin et al, 2012).
Summary of the stem-Gnathostomata
In summary, all the stem gnathostomes were armored fishes, although the nature of the armor - whether consisting of plates or scales - varied through the development of the stem group. Furthermore, the fossil record shows a transition from jawless fishes without teeth to jawed fish with teeth. Other events in the development of the stem group were the appearance of paired fins and articulation between the head and the body.
Transition to the crown group
Some idea of the nature of the transition from the stem group to the crown group of the gnathostomes can be derived from a comparison of the above images with the examples of early crown-Gnathostomata shown below:
Summary of the stem-Gnathostomata
In summary, all the stem gnathostomes were armored fishes, although the nature of the armor - whether consisting of plates or scales - varied through the development of the stem group. Furthermore, the fossil record shows a transition from jawless fishes without teeth to jawed fish with teeth. Other events in the development of the stem group were the appearance of paired fins and articulation between the head and the body.
Transition to the crown group
Some idea of the nature of the transition from the stem group to the crown group of the gnathostomes can be derived from a comparison of the above images with the examples of early crown-Gnathostomata shown below:

Names in red indicate that the fossil is younger than the oldest known crown-group fossil.
Figure 9. Examples of early crown-Gnathostomata
It is noteworthy that the crown gnathostomes shown above, which appear quite similar to modern fishes, are significantly different in general appearance, according to the life restorations shown above, from the Thelodonti and many other stem gnathostomes.
Time frame
Making allowance for the ghost lineage on the gnathostome stem line (Figure 1.1), the time between the origin of the gnathostome stem group and the initiation of the crown group was around 61 million years. Most of the known stem-group fossils, those with red labels in the above figures, are younger than the crown group; there was obviously an explosion of new genera from the Early Silurian to the Early Devonian, but this must have been derived from ancestors that separated from the stem line much earlier, as indicated in the time tree (see Figure 1.2).
Time frame
Making allowance for the ghost lineage on the gnathostome stem line (Figure 1.1), the time between the origin of the gnathostome stem group and the initiation of the crown group was around 61 million years. Most of the known stem-group fossils, those with red labels in the above figures, are younger than the crown group; there was obviously an explosion of new genera from the Early Silurian to the Early Devonian, but this must have been derived from ancestors that separated from the stem line much earlier, as indicated in the time tree (see Figure 1.2).
References
Benton, M. J. (2015). Vertebrate Palaeontology - Fourth edition. John Wiley & Sons, 468 pages.
Benton, M. J., Donoghue, P. C., Asher, R. J., Friedman, M., Near, T. J., & Vinther, J. (2015). Constraints on the timescale of animal evolutionary history. Palaeontologia Electronica, 18(1), 1-106.
Brazeau, M. D., & Friedman, M. (2014). The characters of Palaeozoic jawed vertebrates. Zoological journal of the Linnean Society, 170(4), 779-821.
Donoghue, P. C., & Smith, M. P. (2001). The anatomy of Turinia pagei (Powrie), and the phylogenetic status of the Thelodonti. Earth and Environmental Science Transactions of The Royal Society of Edinburgh, 92(1), 15-37.
Gagnier, P. Y., Blieck, A. R., & Rodrigo, G. (1986). First Ordovician vertebrate from South America. Geobios, 19(5), 629-634.
Giles, S., Friedman, M., & Brazeau, M. D. (2015). Osteichthyan-like cranial conditions in an Early Devonian stem gnathostome. Nature, 520(7545), 82-85.
Janvier, P. (1984). The relationships of the Osteostraci and Galeaspida. Journal of Vertebrate Paleontology, 4(3), 344-358.
Janvier, P. (1987). The paired fins of anaspids: one more hypothesis about their function. Journal of Paleontology, 61(4), 850-853.
Jobbins, M., Rücklin, M., Argyriou, T., & Klug, C. (2021). A large Middle Devonian eubrachythoracid ‘placoderm’(Arthrodira) jaw from northern Gondwana. Swiss Journal of Palaeontology, 140, 1-17.
Johanson, Z., & Smith, M. M. (2005). Origin and evolution of gnathostome dentitions: a question of teeth and pharyngeal denticles in placoderms. Biological Reviews, 80(2), 303-345.
Keating, J. N., Marquart, C. L., Marone, F., & Donoghue, P. C. (2018). The nature of aspidin and the evolutionary origin of bone. Nature ecology & evolution, 2(9), 1501-1506.
Ritchie, A., & Gilbert-Tomlinson, J. (1977). First Ordovician vertebrates from the southern hemisphere. Alcheringa, 1(4), 351-368.
Rücklin, M., Donoghue, P. C., Johanson, Z., Trinajstic, K., Marone, F., & Stampanoni, M. (2012). Development of teeth and jaws in the earliest jawed vertebrates. Nature, 491(7426), 748-751.
Sallan, L., Friedman, M., Sansom, R. S., Bird, C. M., & Sansom, I. J. (2018). The nearshore cradle of early vertebrate diversification. Science, 362(6413), 460-464.
Zhu, M., Yu, X., Choo, B., Wang, J., & Jia, L. (2012). An antiarch placoderm shows that pelvic girdles arose at the root of jawed vertebrates. Biology Letters, 8(3), 453-456.
Zhu, M., Ahlberg, P. E., Pan, Z., Zhu, Y., Qiao, T., Zhao, W., ... & Lu, J. (2016). A Silurian maxillate placoderm illuminates jaw evolution. Science, 354(6310), 334-336.
Benton, M. J., Donoghue, P. C., Asher, R. J., Friedman, M., Near, T. J., & Vinther, J. (2015). Constraints on the timescale of animal evolutionary history. Palaeontologia Electronica, 18(1), 1-106.
Brazeau, M. D., & Friedman, M. (2014). The characters of Palaeozoic jawed vertebrates. Zoological journal of the Linnean Society, 170(4), 779-821.
Donoghue, P. C., & Smith, M. P. (2001). The anatomy of Turinia pagei (Powrie), and the phylogenetic status of the Thelodonti. Earth and Environmental Science Transactions of The Royal Society of Edinburgh, 92(1), 15-37.
Gagnier, P. Y., Blieck, A. R., & Rodrigo, G. (1986). First Ordovician vertebrate from South America. Geobios, 19(5), 629-634.
Giles, S., Friedman, M., & Brazeau, M. D. (2015). Osteichthyan-like cranial conditions in an Early Devonian stem gnathostome. Nature, 520(7545), 82-85.
Janvier, P. (1984). The relationships of the Osteostraci and Galeaspida. Journal of Vertebrate Paleontology, 4(3), 344-358.
Janvier, P. (1987). The paired fins of anaspids: one more hypothesis about their function. Journal of Paleontology, 61(4), 850-853.
Jobbins, M., Rücklin, M., Argyriou, T., & Klug, C. (2021). A large Middle Devonian eubrachythoracid ‘placoderm’(Arthrodira) jaw from northern Gondwana. Swiss Journal of Palaeontology, 140, 1-17.
Johanson, Z., & Smith, M. M. (2005). Origin and evolution of gnathostome dentitions: a question of teeth and pharyngeal denticles in placoderms. Biological Reviews, 80(2), 303-345.
Keating, J. N., Marquart, C. L., Marone, F., & Donoghue, P. C. (2018). The nature of aspidin and the evolutionary origin of bone. Nature ecology & evolution, 2(9), 1501-1506.
Ritchie, A., & Gilbert-Tomlinson, J. (1977). First Ordovician vertebrates from the southern hemisphere. Alcheringa, 1(4), 351-368.
Rücklin, M., Donoghue, P. C., Johanson, Z., Trinajstic, K., Marone, F., & Stampanoni, M. (2012). Development of teeth and jaws in the earliest jawed vertebrates. Nature, 491(7426), 748-751.
Sallan, L., Friedman, M., Sansom, R. S., Bird, C. M., & Sansom, I. J. (2018). The nearshore cradle of early vertebrate diversification. Science, 362(6413), 460-464.
Zhu, M., Yu, X., Choo, B., Wang, J., & Jia, L. (2012). An antiarch placoderm shows that pelvic girdles arose at the root of jawed vertebrates. Biology Letters, 8(3), 453-456.
Zhu, M., Ahlberg, P. E., Pan, Z., Zhu, Y., Qiao, T., Zhao, W., ... & Lu, J. (2016). A Silurian maxillate placoderm illuminates jaw evolution. Science, 354(6310), 334-336.
Image credits - Stem-Gnathostomes
- Figure 2 (Sacabambaspis janvieri, fossil): Ghedoghedo / CC BY-SA (https://creativecommons.org/licenses/by-sa/3.0)
- Figure 2 (Sacabambaspis janvieri, life restoration): Nobu Tamura, licensed under Creative Commons Attribution- ShareAlike (CC BY-SA)
- Figure 2 (Arandaspis prionotolepis): Nobu Tamura (http://spinops.blogspot.com) [CC BY-SA 3.0 (https://creativecommons.org/licenses/by-sa/3.0) or GFDL (http://www.gnu.org/copyleft/fdl.html)], from Wikimedia Commons
- Figure 2 (Astraspis desiderata): Philippe Janvier / CC BY (https://creativecommons.org/licenses/by/3.0)
- Figure 2 (Athenaegis chattertoni): Nobu Tamura, licensed under Creative Commons Attribution- ShareAlike (CC BY-SA)
- Figure 2 (Tolypelepis undulata): Apokryltaros / CC BY-SA (https://creativecommons.org/licenses/by-sa/4.0)
- Figure 3 (Jamoytius kerwoodi): Nobu Tamura, licensed under Creative Commons Attribution- ShareAlike (CC BY-SA)
- Figure 3 (Lasanius problematicus): Nobu Tamura, licensed under Creative Commons Attribution- ShareAlike (CC BY-SA)
- Figure 3 (Birkenia elegans, fossil): Ghedoghedo / CC BY-SA (https://creativecommons.org/licenses/by-sa/3.0)
- Figure 3 (Birkenia elegans, life restoration): Nobu Tamura, licensed under Creative Commons Attribution- ShareAlike (CC BY-SA)
- Figure 4 (Archipelepis turbinata): Nobu Tamura, licensed under Creative Commons Attribution- ShareAlike (CC BY-SA)
- Figure 4 (Pezopallichthys ritchiei): Apokryltaros / CC BY-SA (https://creativecommons.org/licenses/by-sa/4.0)
- Figure 4 (Loganellia scotica, fossil): Ghedoghedo / CC BY-SA (https://creativecommons.org/licenses/by-sa/4.0)
- Figure 4 (Loganellia scotica, life restoration): Nobu Tamura, licensed under Creative Commons Attribution- ShareAlike (CC BY-SA)
- Figure 4 (Pituriaspis doylei): Nobu Tamura, licensed under Creative Commons Attribution- ShareAlike (CC BY-SA)
- Figure 4 (Neeyambaspis enigmatica): Apokryltaros at English Wikipedia, CC BY-SA 3.0 <http://creativecommons.org/licenses/by-sa/3.0/>, via Wikimedia Commons
- Figure 5 (Hanyangaspis guodingshanensis): Nobu Tamura, licensed under Creative Commons Attribution- ShareAlike (CC BY-SA)
- Figure 5 (Dayongaspis hunanensis): Apokryltaros / CC BY-SA (https://creativecommons.org/licenses/by-sa/4.0)
- Figure 6 (Ateleaspis tesselata): Nobu Tamura, licensed under Creative Commons Attribution- ShareAlike (CC BY-SA)
- Figure 6 (Hemicyclaspis murchisoni, fossil): Ghedoghedo / CC BY-SA (https://creativecommons.org/licenses/by-sa/3.0)
- Figure 6 (Hemicyclaspis murchisoni, life restoration): Nobu Tamura, licensed under Creative Commons Attribution- ShareAlike (CC BY-SA)
- Figure 7 (Shimenolepis graniferus): Apokryltaros / CC BY-SA (https://creativecommons.org/licenses/by-sa/3.0)
- Figure 7 (Silurolepis platydorsalis): Apokryltaros / CC BY-SA (https://creativecommons.org/licenses/by-sa/3.0)
- Figure 7 (Yunnanolepis chii): Nobu Tamura, licensed under Creative Commons Attribution- ShareAlike (CC BY-SA)
- Figure 7 (Zhanjilepis aspratilis): Apokryltaros / CC BY-SA (https://creativecommons.org/licenses/by-sa/3.0)
- Figure 7 (Chuchinolepis dongmoensis): Apokryltaros, CC BY-SA 3.0 <https://creativecommons.org/licenses/by-sa/3.0>, via Wikimedia Commons
- Figure 7 (Minicrania lirouyii): Apokryltaros / CC BY-SA (https://creativecommons.org/licenses/by-sa/3.0)
- Figure 8 (Palaeacanthaspis vasta): Apokryltaros / CC BY-SA (https://creativecommons.org/licenses/by-sa/3.0)
- Figure 8 (Diandongpetalichthys liaojiaoshanensis): Apokryltaros / CC BY-SA (https://creativecommons.org/licenses/by-sa/3.0)
- Figure 8 (Romundina stellina): Apokryltaros / CC BY-SA (https://creativecommons.org/licenses/by-sa/3.0)
- Figure 8 (Kujdanowiaspis sp.): Entelognathus, CC BY-SA 4.0 <https://creativecommons.org/licenses/by-sa/4.0>, via Wikimedia Commons
- Figure 8 (Dicksonosteus arcticus): Entelognathus, CC BY-SA 4.0 <https://creativecommons.org/licenses/by-sa/4.0>, via Wikimedia Commons
- Figure 8 (Coccosteus cuspidatus, fossil): Ghedoghedo / CC BY-SA (https://creativecommons.org/licenses/by-sa/3.0)
- Figure 8 (Coccosteus cuspidatus, life restoration): Estonian Museum of Natural History / CC BY-SA (https://creativecommons.org/licenses/by-sa/4.0)
- Figure 8 (Buchanosteus confertituberculatus): Apokryltaros / CC BY-SA (https://creativecommons.org/licenses/by-sa/3.0)
- Figure 8 (Qilinyu rostrata): Apokryltaros / CC BY-SA (https://creativecommons.org/licenses/by-sa/4.0)
- Figure 8 (Entelognathus primordialis): Nobu Tamura, licensed under Creative Commons Attribution- ShareAlike (CC BY-SA)
- Figure 9 (Nerepisacanthus denisoni): Danielle Dufault [CC BY 2.5 (https://creativecommons.org/licenses/by/2.5)]
- Figure 9 (Parexus recurvus): Nobu Tamura, licensed under Creative Commons Attribution- ShareAlike (CC BY-SA)
- Figure 9 ('Ligulalepis' ): Open Access article Clement, A. M., King, B., Giles, S., Choo, B., Ahlberg, P. E., Young, G. C., & Long, J. A. (2018). Neurocranial anatomy of an enigmatic Early Devonian fish sheds light on early osteichthyan evolution. Elife, 7, e34349.