Introduction
The appearance of the eukaryotes (all organisms that have cells in which the nucleus is enclosed within a membrane) represents one of the most fundamental milestones in the history of life (Butterfield, 2015). This transition occurred when the eukaryotes split off from the prokaryotic Archaea by acquiring the intra-cellular organization and other features that distinguishes them from the prokaryotes. The process by which this transformation occurred remains unclear (Galbaldón and Pittis, 2015).
In order to place the appearance of eukaryotes in the context of geological time, we need to be able to find evidence of their existence in the rocks. Molecular biomarkers and isotopes have been invoked, but for the eukaryotes do not provide evidence for an earlier origin than do physical fossils (Javaux and Lepot, 2018).
Recognition of eukaryotes in the fossil record requires recognition of the synapomorphies associated with the Eukaryota. Unfortunately, the known synapomorphies of the eukaryotes are defined almost exclusively in terms of what can be observed inside extant cells (Fehling et al, 2007). Detecting those features in ancient fossils is extremely difficult or impossible. For this reason, other characteristics have been suggested, such as large size (40-250μm) and complex wall structure including striations, longitudinal ruptures, and a trilaminar organization (Betts et al, 2018). Porter (2020) states that “the oldest widely accepted evidence of eukaryotes is large (greater than 100 µm), spiny, ornamented, organic walled microfossils found in latest Paleoproterozoic rocks (ca 1650 Ma)”. This description is reminiscent of the definition of organisms known as “acritarchs” according to Evitt (1963):
Small microfossils of unknown and probably varied biological affinities consisting of a central cavity enclosed by a wall of single or multiple layers and of chiefly organic composition; symmetry, shape, structure, and ornamentation varied; central cavity closed or communicating with the exterior by varied means, for example: pores, a slitlike or irregular rupture, a circular opening (the pylome).
Now this definition does not allow all acritarchs to be identified exclusively as eukaryotes, as some organisms previously classed as acritarchs have since been placed in the prokaryotic cyanobacteria (Nowak et al, 2015). However, there is evidence that eukaryotic acritarchs existed as far back as the Proterozoic (Baludikay et al, 2015).
These considerations indicate that it is possible to recognize eukaryotic microfossils in the fossil record. However, it is not possible to say whether they belong to the stem group or the crown group, and so they can be considered only as members of the total group of the Eukaryota (Betts et al, 2018).
In order to place the appearance of eukaryotes in the context of geological time, we need to be able to find evidence of their existence in the rocks. Molecular biomarkers and isotopes have been invoked, but for the eukaryotes do not provide evidence for an earlier origin than do physical fossils (Javaux and Lepot, 2018).
Recognition of eukaryotes in the fossil record requires recognition of the synapomorphies associated with the Eukaryota. Unfortunately, the known synapomorphies of the eukaryotes are defined almost exclusively in terms of what can be observed inside extant cells (Fehling et al, 2007). Detecting those features in ancient fossils is extremely difficult or impossible. For this reason, other characteristics have been suggested, such as large size (40-250μm) and complex wall structure including striations, longitudinal ruptures, and a trilaminar organization (Betts et al, 2018). Porter (2020) states that “the oldest widely accepted evidence of eukaryotes is large (greater than 100 µm), spiny, ornamented, organic walled microfossils found in latest Paleoproterozoic rocks (ca 1650 Ma)”. This description is reminiscent of the definition of organisms known as “acritarchs” according to Evitt (1963):
Small microfossils of unknown and probably varied biological affinities consisting of a central cavity enclosed by a wall of single or multiple layers and of chiefly organic composition; symmetry, shape, structure, and ornamentation varied; central cavity closed or communicating with the exterior by varied means, for example: pores, a slitlike or irregular rupture, a circular opening (the pylome).
Now this definition does not allow all acritarchs to be identified exclusively as eukaryotes, as some organisms previously classed as acritarchs have since been placed in the prokaryotic cyanobacteria (Nowak et al, 2015). However, there is evidence that eukaryotic acritarchs existed as far back as the Proterozoic (Baludikay et al, 2015).
These considerations indicate that it is possible to recognize eukaryotic microfossils in the fossil record. However, it is not possible to say whether they belong to the stem group or the crown group, and so they can be considered only as members of the total group of the Eukaryota (Betts et al, 2018).
Total-Eukaryota
The oldest known members of the eukaryote total group are organic-walled microfossils (acritarchs) from the late Paleoproterozoic (Statherian) Changcheng Group in the Yanshan Range, North China (Miao et al, 2019). Some of these are illustrated below (click on image for larger view):

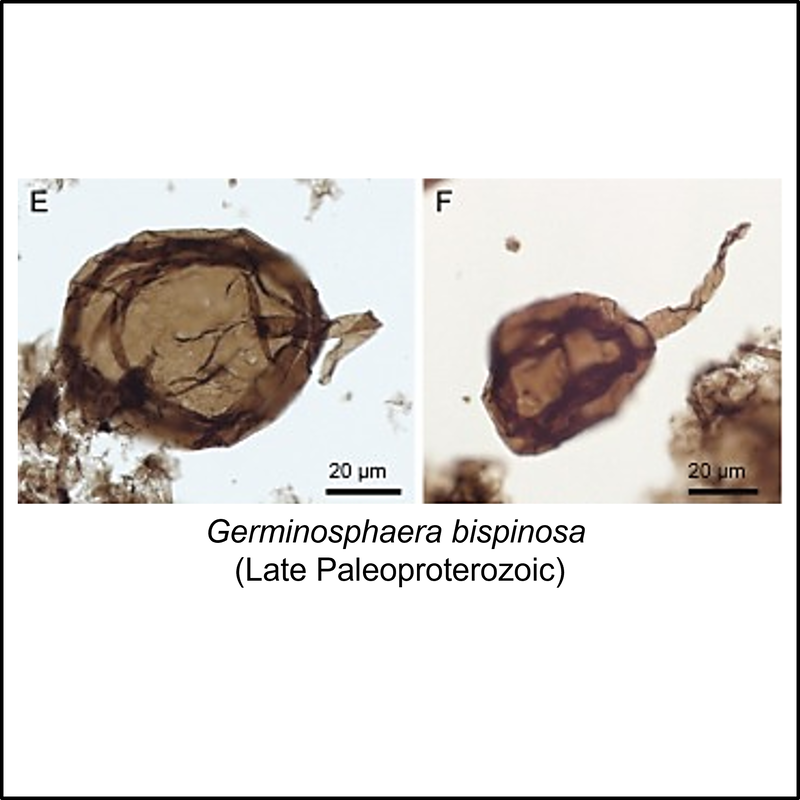
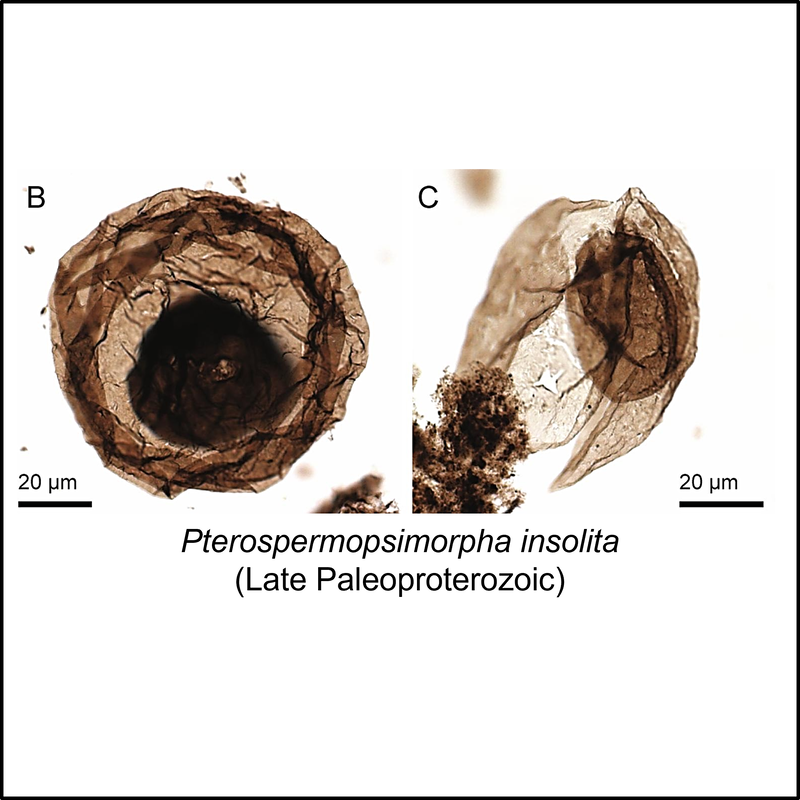
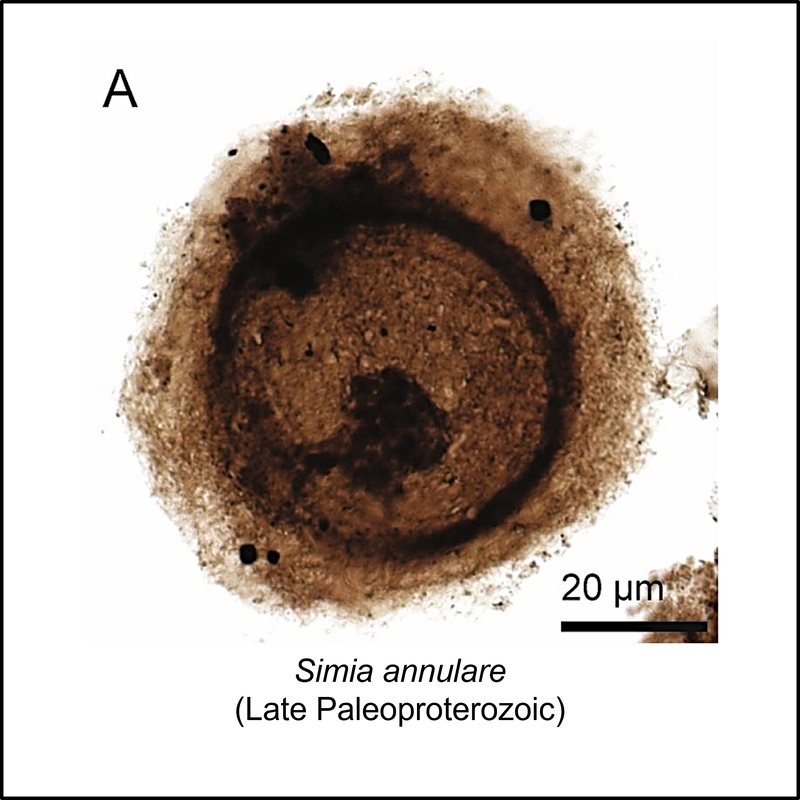

Crown-Eukaryota
The oldest fossil that can be confidently assigned to the crown group of the eukaryotes is Bangiomorpha pubescens, a red alga (total-Rhodophyta) from the late Mesoproterozoic (Stenian) Lower Hunting Formation, of Somerset Island, Arctic Canada (Gibson et al, 2018). The only image available in the public domain is shown below:
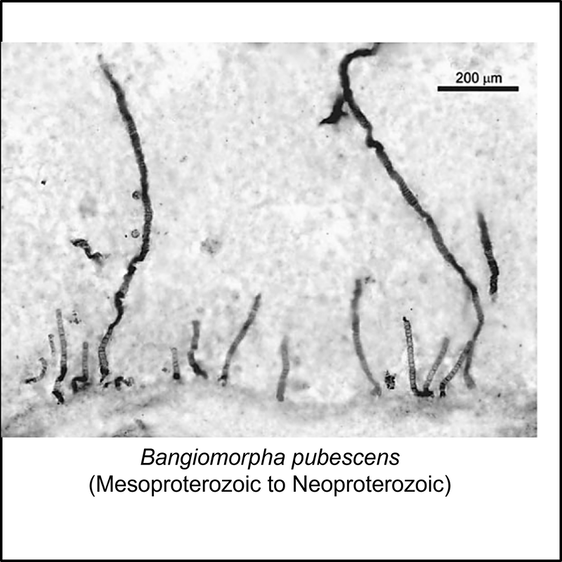
Comparison of the ages of the oldest total-group eukaryotes and Bangiomorpha pubescens indicates that there is a period of uncertainty of at least 564 million years (between 1,624 and 1,060 million years ago) for the time of appearance of the eukaryote crown group.
References
Baludikay, B. K., Storme, J. Y., François, C., Baudet, D., & Javaux, E. J. (2016). A diverse and exquisitely preserved organic-walled microfossil assemblage from the Meso–Neoproterozoic Mbuji-Mayi Supergroup (Democratic Republic of Congo) and implications for Proterozoic biostratigraphy. Precambrian research, 281, 166-184.
Betts, H. C., Puttick, M. N., Clark, J. W., Williams, T. A., Donoghue, P. C., & Pisani, D. (2018). Integrated genomic and fossil evidence illuminates life’s early evolution and eukaryote origin. Nature ecology & evolution, 2(10), 1556-1562.
Butterfield, N. J. (2015). Early evolution of the Eukaryota. Palaeontology, 58(1), 5-17.
Evitt, W. R. (1963). A DISCUSSION AND PROPOSALS CONCERNING FOSSIL DINOFLAGELLATES, HYSTRICHOSPHERES, AND ACRITARCHS, II. Proceedings of the National Academy of Sciences of the United States of America, 49(3), 298-302.
Fehling, J., Stoeker, D., & Baldauf, S. L. (2007). Photosynthesis and the eukaryote tree of life. In Evolution of primary producers in the sea (pp. 75-107). Academic Press.
Gabaldón, T., & Pittis, A. A. (2015). Origin and evolution of metabolic sub-cellular compartmentalization in eukaryotes. Biochimie, 119, 262-268.
Gibson, T. M., Shih, P. M., Cumming, V. M., Fischer, W. W., Crockford, P. W., Hodgskiss, M. S., ... & Halverson, G. P. (2018). Precise age of Bangiomorpha pubescens dates the origin of eukaryotic photosynthesis. Geology, 46(2), 135-138.
Javaux, E. J., & Lepot, K. (2018). The Paleoproterozoic fossil record: implications for the evolution of the biosphere during Earth's middle-age. Earth-Science Reviews, 176, 68-86.
Miao, L., Moczydłowska, M., Zhu, S., & Zhu, M. (2019). New record of organic-walled, morphologically distinct microfossils from the late Paleoproterozoic Changcheng Group in the Yanshan Range, North China. Precambrian Research, 321, 172-198.
Nowak, H., Servais, T., Monnet, C., Molyneux, S. G., & Vandenbroucke, T. R. (2015). Phytoplankton dynamics from the Cambrian Explosion to the onset of the Great Ordovician Biodiversification Event: a review of Cambrian acritarch diversity. Earth-Science Reviews, 151, 117-131.
Porter, S. M. (2020). Insights into eukaryogenesis from the fossil record. Interface Focus, 10(4), 20190105.
Betts, H. C., Puttick, M. N., Clark, J. W., Williams, T. A., Donoghue, P. C., & Pisani, D. (2018). Integrated genomic and fossil evidence illuminates life’s early evolution and eukaryote origin. Nature ecology & evolution, 2(10), 1556-1562.
Butterfield, N. J. (2015). Early evolution of the Eukaryota. Palaeontology, 58(1), 5-17.
Evitt, W. R. (1963). A DISCUSSION AND PROPOSALS CONCERNING FOSSIL DINOFLAGELLATES, HYSTRICHOSPHERES, AND ACRITARCHS, II. Proceedings of the National Academy of Sciences of the United States of America, 49(3), 298-302.
Fehling, J., Stoeker, D., & Baldauf, S. L. (2007). Photosynthesis and the eukaryote tree of life. In Evolution of primary producers in the sea (pp. 75-107). Academic Press.
Gabaldón, T., & Pittis, A. A. (2015). Origin and evolution of metabolic sub-cellular compartmentalization in eukaryotes. Biochimie, 119, 262-268.
Gibson, T. M., Shih, P. M., Cumming, V. M., Fischer, W. W., Crockford, P. W., Hodgskiss, M. S., ... & Halverson, G. P. (2018). Precise age of Bangiomorpha pubescens dates the origin of eukaryotic photosynthesis. Geology, 46(2), 135-138.
Javaux, E. J., & Lepot, K. (2018). The Paleoproterozoic fossil record: implications for the evolution of the biosphere during Earth's middle-age. Earth-Science Reviews, 176, 68-86.
Miao, L., Moczydłowska, M., Zhu, S., & Zhu, M. (2019). New record of organic-walled, morphologically distinct microfossils from the late Paleoproterozoic Changcheng Group in the Yanshan Range, North China. Precambrian Research, 321, 172-198.
Nowak, H., Servais, T., Monnet, C., Molyneux, S. G., & Vandenbroucke, T. R. (2015). Phytoplankton dynamics from the Cambrian Explosion to the onset of the Great Ordovician Biodiversification Event: a review of Cambrian acritarch diversity. Earth-Science Reviews, 151, 117-131.
Porter, S. M. (2020). Insights into eukaryogenesis from the fossil record. Interface Focus, 10(4), 20190105.
Image credits - Eukaryotes
- Header: Amanita muscaria toadstool By Onderwijsgek at nl.wikipedia / CC BY-SA 3.0 NL (https://creativecommons.org/licenses/by-sa/3.0/nl/deed.en)
- All except Bangiomorpha pubescens are from Open Access article Loron, C. C., Rainbird, R. H., Turner, E. C., Greenman, J. W., & Javaux, E. J. (2019). Organic-walled microfossils from the late Mesoproterozoic to early Neoproterozoic lower Shaler Supergroup (Arctic Canada): diversity and biostratigraphic significance. Precambrian Research, 321, 349-374.
- Bangiomorpha pubescens: from Open Access article Garwood, R. J. (2012). Patterns In Palaeontology: The first 3 billion years of evolution. Palaeontology Online, 2(11), 1-14.