This page deals with the stem group of the ray-finned fishes (Superclass Actinopterygii, clade Osteichthyes). The actinopterygians are a very diverse group, consisting of some 42 orders containing more than 480 families, but share the characteristic of having fins that are supported by rays of dermal bone rather than by cartilage (Encyclopedia Britannica).
Many stem-group actinopterygian fossils have been found. Their phylogenetic relationships have been elucidated in many morphological analyses (e.g. Argyriou et al (2018), Wilson et al (2018), Ren and Xu (2021)), of which that by Giles et al (2017), shown in the phylogenetic time tree below, is a representative example:
Many stem-group actinopterygian fossils have been found. Their phylogenetic relationships have been elucidated in many morphological analyses (e.g. Argyriou et al (2018), Wilson et al (2018), Ren and Xu (2021)), of which that by Giles et al (2017), shown in the phylogenetic time tree below, is a representative example:
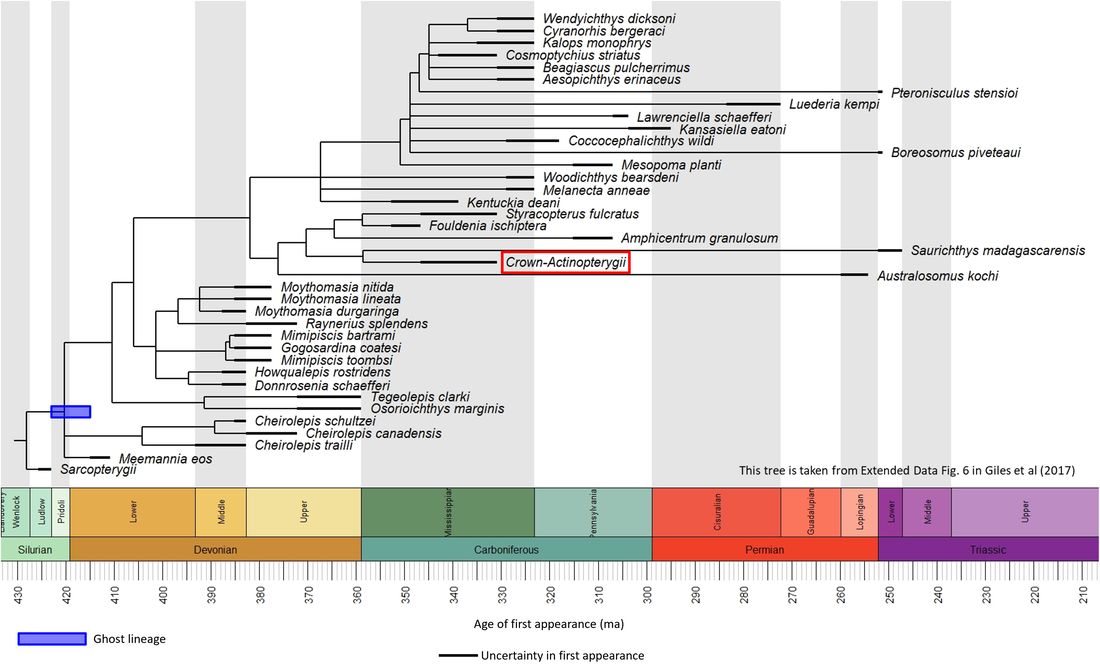
Figure 1. Time tree of the stem-Actinopterygii
The oldest known member of the stem-Actinopterygii is Meemannia eos, described from the middle part of the Xitun Formation, of Early Devonian (Late Lochkovian) age, at a locality close to Xitun village in the suburb of Qujing, Yunnan, southwestern China (Zhu et al, 2010; Giles et al, 2017). No images of this species are available in the public domain. However, the other stem-group fossils for which public domain images are available (unfortunately rather few relative to the number of species shown in Figure 1 above) are shown below (click on image for a larger view):
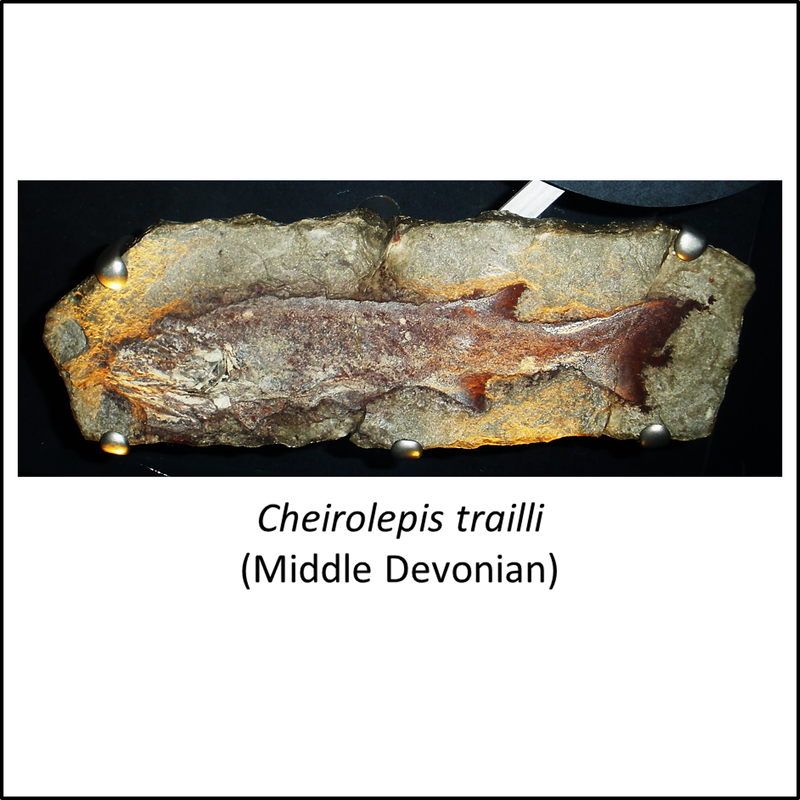



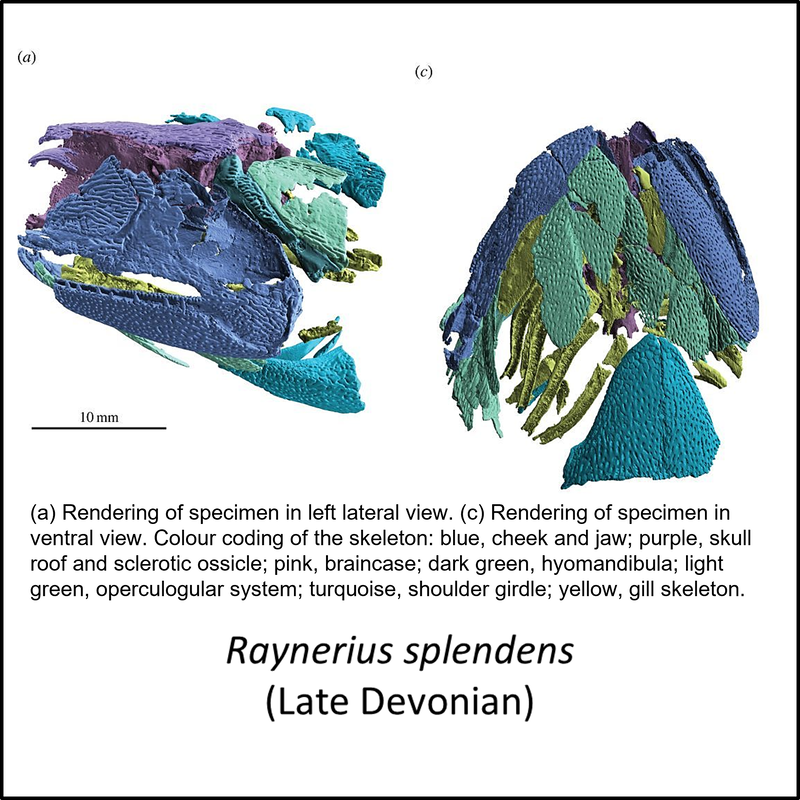







Names in red indicate that the fossil is younger than the oldest known crown-group fossil.
Figure 2. Images of stem-group Actinopterygii, with the crown-group example Platysomus sp.
The images shown above are placed in left-to-right order from most basal towards the crown group, but no obvious trends can be seen.
Some idea of the nature of the transition from the stem group to the crown group of the actinopterygian can be derived from a comparison of the above images with the examples of early crown-Actinopterygii shown below:
Some idea of the nature of the transition from the stem group to the crown group of the actinopterygian can be derived from a comparison of the above images with the examples of early crown-Actinopterygii shown below:

Figure 3. Examples of early crown-Actinopterygii
The actinopterygian stem line includes a ghost lineage (shown as a blue bar in Figure 1), because the oldest known stem-group sarcopterygian (of Late Silurian age; see page on the Sarcopterygii) is somewhat older than the stem-group Actinopterygii, which are of Early Devonian (Lochkovian) and younger age. Given that the two stem groups must have appeared at the same time, the actinopterygian stem-group transition must also have begun in the Late Silurian. Comparing this age with that of the oldest member of the crown-Actinopterygii indicates that the stem-to-crown transition lasted at least 76 million years, from the Late Silurian to the Early Carboniferous (Figure 1). A histogram of the rates of appearance of the known stem-group genera is shown below:

Figure 4. Rate of appearance of stem-Actinopterygii (predating the crown group and including only the genera shown in Figure 1)
References
Argyriou, T., Giles, S., Friedman, M., Romano, C., Kogan, I., & Sánchez-Villagra, M. R. (2018). Internal cranial anatomy of Early Triassic species of †Saurichthys (Actinopterygii: †Saurichthyiformes): implications for the phylogenetic placement of †Saurichthyiforms. BMC evolutionary biology, 18(1), 1-41.
Dineley, D. & Metcalf, S. (1999). Fossil Fishes of Great Britain, Geological Conservation Review Series, No. 16, Joint Nature Conservation Committee, Peterborough, 675 pp.
Giles, S., Xu, G. H., Near, T. J., & Friedman, M. (2017). Early members of ‘living fossil’ lineage imply later origin of modern ray-finned fishes. Nature, 549(7671), 265-268.
Lu, J., Giles, S., Friedman, M., den Blaauwen, J. L., & Zhu, M. (2016). The oldest actinopterygian highlights the cryptic early history of the hyperdiverse ray-finned fishes. Current Biology, 26(12), 1602-1608.
Ren, Y., & Xu, G. H. (2021). A new species of Pteronisculus from the Middle Triassic (Anisian) of Luoping, Yunnan, China, and phylogenetic relationships of early actinopterygian fishes. Vertebrata PalAsiatica, 59(3), 169-199.
Wilson, C. D., Pardo, J. D., & Anderson, J. S. (2018). A primitive actinopterygian braincase from the Tournaisian of Nova Scotia. Royal Society open science, 5(5), 171727.
Zhu, M., Wang, W., & Yu, X. (2010). Meemannia eos, a basal sarcopterygian fish from the Lower Devonian of China–expanded description and significance. Morphology, phylogeny and paleobiogeography of fossil fishes. München: Verlag Dr. Friedrich Pfeil, 199-214.
Dineley, D. & Metcalf, S. (1999). Fossil Fishes of Great Britain, Geological Conservation Review Series, No. 16, Joint Nature Conservation Committee, Peterborough, 675 pp.
Giles, S., Xu, G. H., Near, T. J., & Friedman, M. (2017). Early members of ‘living fossil’ lineage imply later origin of modern ray-finned fishes. Nature, 549(7671), 265-268.
Lu, J., Giles, S., Friedman, M., den Blaauwen, J. L., & Zhu, M. (2016). The oldest actinopterygian highlights the cryptic early history of the hyperdiverse ray-finned fishes. Current Biology, 26(12), 1602-1608.
Ren, Y., & Xu, G. H. (2021). A new species of Pteronisculus from the Middle Triassic (Anisian) of Luoping, Yunnan, China, and phylogenetic relationships of early actinopterygian fishes. Vertebrata PalAsiatica, 59(3), 169-199.
Wilson, C. D., Pardo, J. D., & Anderson, J. S. (2018). A primitive actinopterygian braincase from the Tournaisian of Nova Scotia. Royal Society open science, 5(5), 171727.
Zhu, M., Wang, W., & Yu, X. (2010). Meemannia eos, a basal sarcopterygian fish from the Lower Devonian of China–expanded description and significance. Morphology, phylogeny and paleobiogeography of fossil fishes. München: Verlag Dr. Friedrich Pfeil, 199-214.
Image credits – Stem-Actinopterygii
- Figure 2 (Cheirolepis trailli, fossil): Ghedoghedo, CC BY-SA 3.0 <https://creativecommons.org/licenses/by-sa/3.0>, via Wikimedia Commons
- Figure 2 (Cheirolepis trailli, life restoration): Nobu Tamura under a Creative Commons 3.0 Unported (CC BY-NC-ND 3.0) license
- Figure 2 (Cheirolepis canadensis): Placoderm2, CC BY-SA 4.0 <https://creativecommons.org/licenses/by-sa/4.0>, via Wikimedia Commons
- Figure 2 (Howqualepis rostridens): Copyright Museums Victoria / CC BY (Licensed as Attribution 4.0 International)
- Figure 2 (Raynerius splendens): Open Access article Giles, S., Darras, L., Clément, G., Blieck, A., & Friedman, M. (2015). An exceptionally preserved Late Devonian actinopterygian provides a new model for primitive cranial anatomy in ray-finned fishes. Proceedings of the Royal Society B: Biological Sciences, 282(1816), 20151485.
- Figure 2 (Moythomasia nitida): Oilshale, CC BY-SA 4.0 <https://creativecommons.org/licenses/by-sa/4.0>, via Wikimedia Commons
- Figure 2 (Woodichthys bearsdeni): British Geological Survey under a Creative Commons Attribution-NonCommercial-ShareAlike 3.0 Unported License.
- Figure 2 (Boreosomus gillioti): Ghedoghedo [CC BY-SA 3.0 (https://creativecommons.org/licenses/by-sa/3.0)]
- Figure 2 (Cyranorhis bergeraci): Ryan Somma, CC BY-SA 2.0 <https://creativecommons.org/licenses/by-sa/2.0>, via Wikimedia Commons
- Figure 2 (Australosomus merlei): Emőke Dénes, CC BY-SA 4.0 <https://creativecommons.org/licenses/by-sa/4.0>, via Wikimedia Commons
- Figure 2 (Saurichthys sp., fossil): Ghedoghedo, CC BY-SA 4.0 <https://creativecommons.org/licenses/by-sa/4.0>, via Wikimedia Commons
- Figure 2 (Saurichthys sp., life restoration): Bildflut [CC0]
- Figure 3 (Tanaocrossus kalliokoskii): Open Access article Hunt-Foster, R., Lockley, M., Milner, A., Foster, J., Matthews, N., Breithaupt, B., & Smith, J. (2016). Tracking dinosaurs in BLM canyon country, Utah. Geology of the intermountain west, 3, 67-100.
- Figure 3 (Discoserra pectinodon): Gyik Toma (Tommy the paleobear) from Romania, CC BY 2.0 <https://creativecommons.org/licenses/by/2.0>, via Wikimedia Commons
- Figure 3 (Platysomus superbus): Open Access article Wilson, C. D., Mansky, C. F., & Anderson, J. S. (2021). A platysomid occurrence from the Tournaisian of Nova Scotia. Scientific reports, 11(1), 1-12.