This page is the first in a series of pages that covers fossils associated with the stem group that leads to the crown group of the tetrapods (Superclass Tetrapoda, Infraphylum Gnathostomata) and with the stem groups of the clades within the crown group.
A summary of the phylogeny of this clade is shown in the following two-part figure::
A summary of the phylogeny of this clade is shown in the following two-part figure::
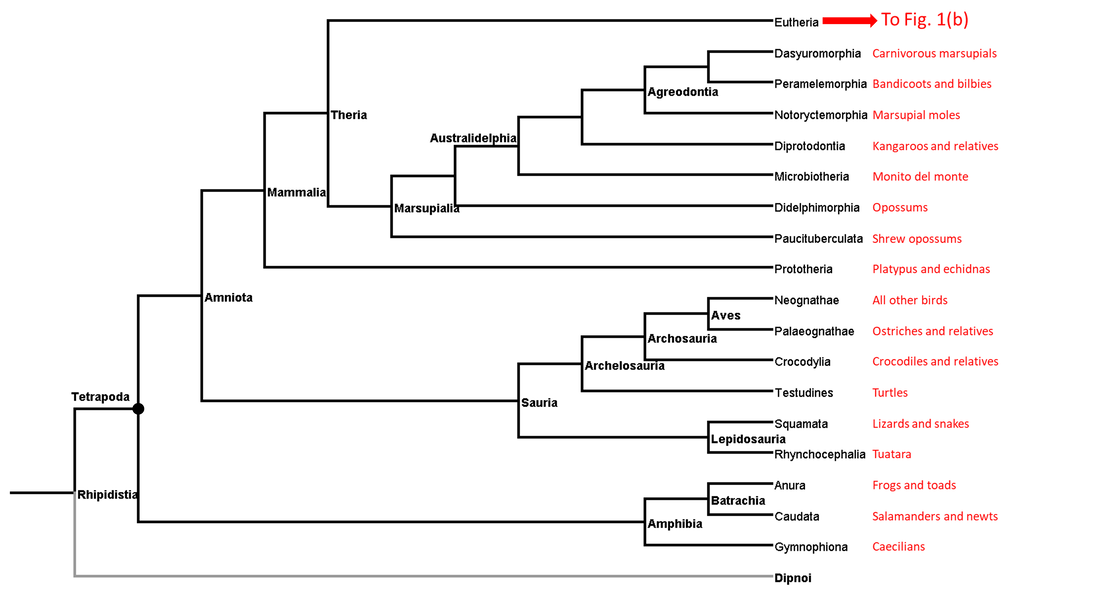
Figure 1a. Summarized phylogenetic tree of the tetrapods (part 1)
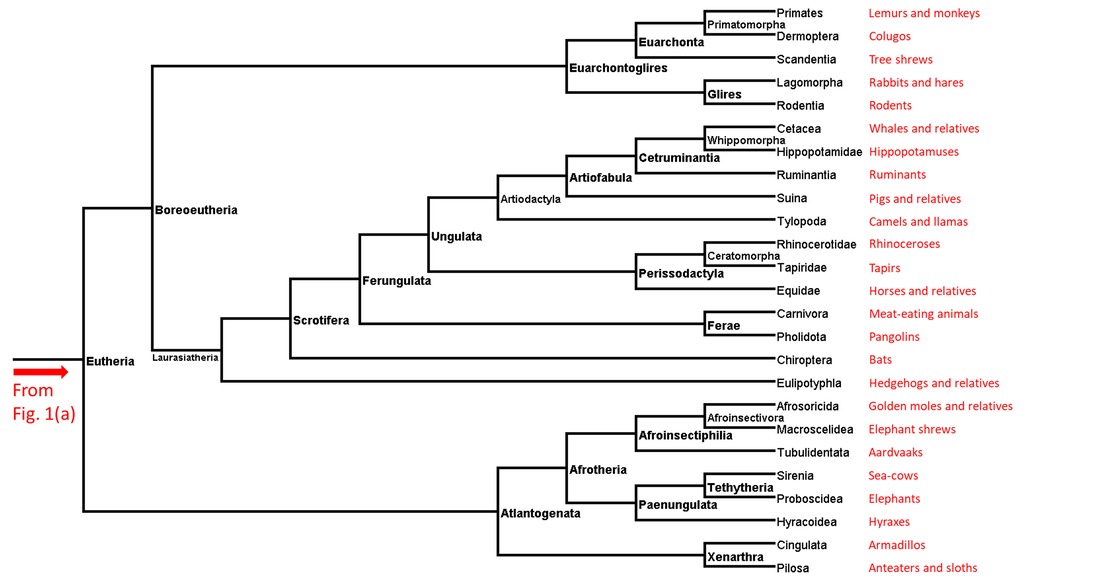
Figure 1b. Summarized phylogenetic tree of the tetrapods (part 2)
Each page of this website deals with a single branch of the phylogenetic tree. Links to the pages corresponding to the branches shown above (those for which adequate data are available) can be found under the Navigation tab above.
The tetrapod stem branches not represented in this website are listed in the following table. In each case the column highlighted by the letter X indicates the reason or reasons for not including that branch:
The tetrapod stem branches not represented in this website are listed in the following table. In each case the column highlighted by the letter X indicates the reason or reasons for not including that branch:

The remainder of this page deals with the stem-group tetrapods.
The evolution of the stem-Tetrapoda is a topic that is undergoing active research at the present time, and new phylogenetic trees continue to be published. Those published since around 2008 are broadly in agreement, although the placement of some fossil species does vary significantly. A representative tree, constructed by combining two recent publications, is shown below:
The evolution of the stem-Tetrapoda is a topic that is undergoing active research at the present time, and new phylogenetic trees continue to be published. Those published since around 2008 are broadly in agreement, although the placement of some fossil species does vary significantly. A representative tree, constructed by combining two recent publications, is shown below:
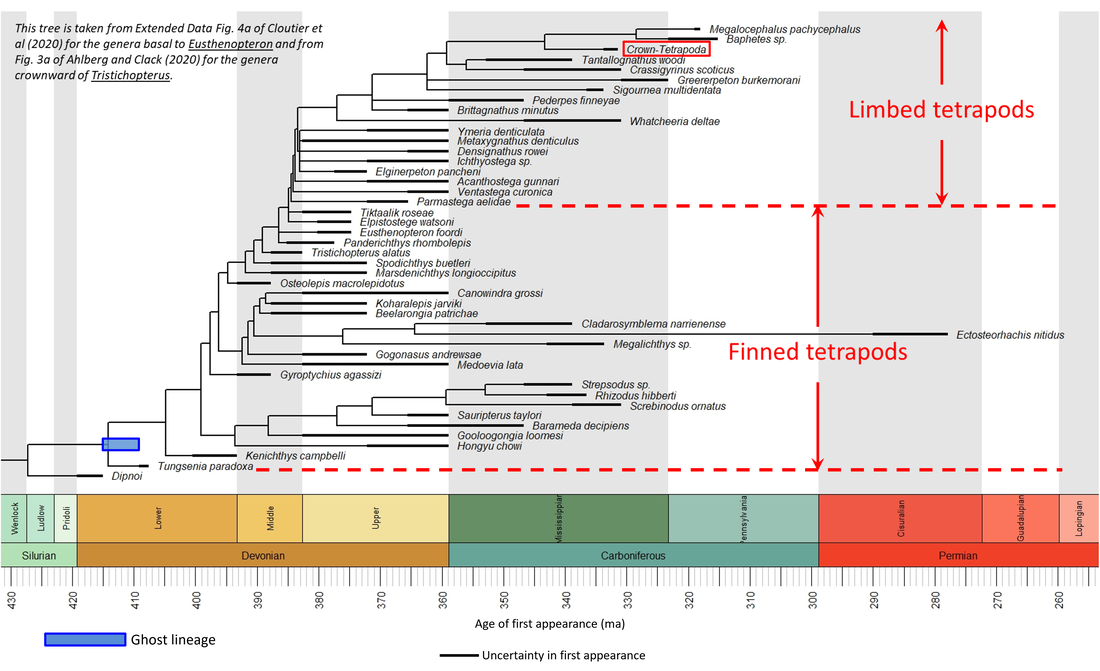
Figure 2. Phylogenetic time tree of the stem-Tetrapoda
The above tree represents a trend from species that are clearly fishes to those that are clearly four-legged animals. There is thus a transition from finned tetrapods to limbed tetrapods (as, for example, in Figure 4 of Hirasawa et al, 2022). There is disagreement about whether Tiktaalik roseae, for example, is a finned (Ruta et al, 2018; Molnar et al, 2020; Hirasawa et al, 2022) or a limbed tetrapod (Dickson et al, 2021), but by all accounts that species is a key representative of the transition from fishes to land animals (Shubin et al, 2006; Hohn-Schulte et al, 2013).
The oldest known member of the stem group is Tungsenia paradoxa, from the Posongchong Formation of Pragian (Early Devonian) age near Zhaotong in northeast Yunnan, China (Lu et al, 2012). A life restoration of this species is shown below, together with other stem-Tetrapoda for which public-domain images are available (click on image to see a larger version):
The oldest known member of the stem group is Tungsenia paradoxa, from the Posongchong Formation of Pragian (Early Devonian) age near Zhaotong in northeast Yunnan, China (Lu et al, 2012). A life restoration of this species is shown below, together with other stem-Tetrapoda for which public-domain images are available (click on image to see a larger version):















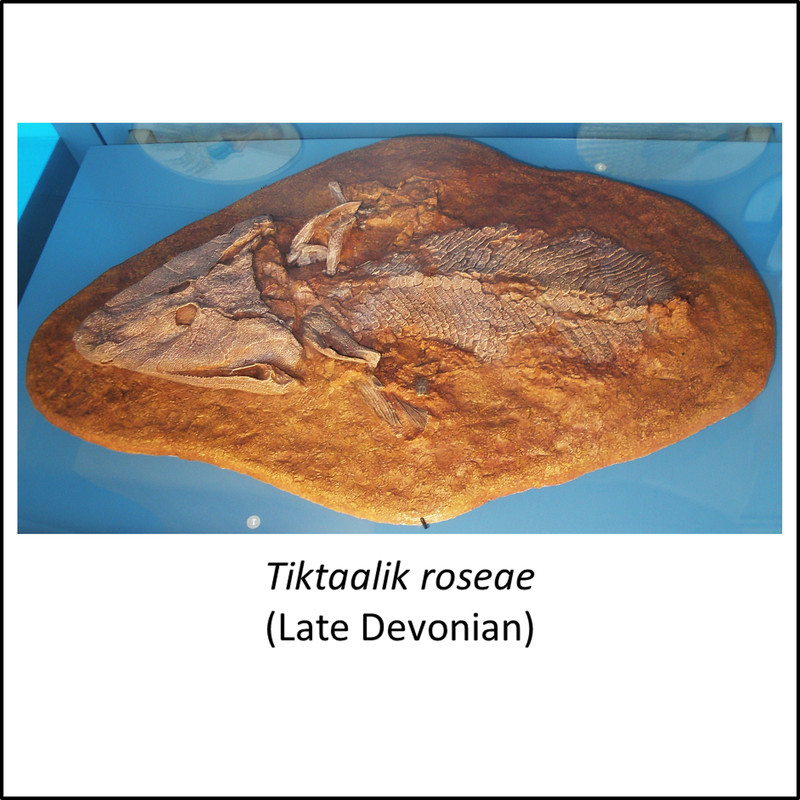

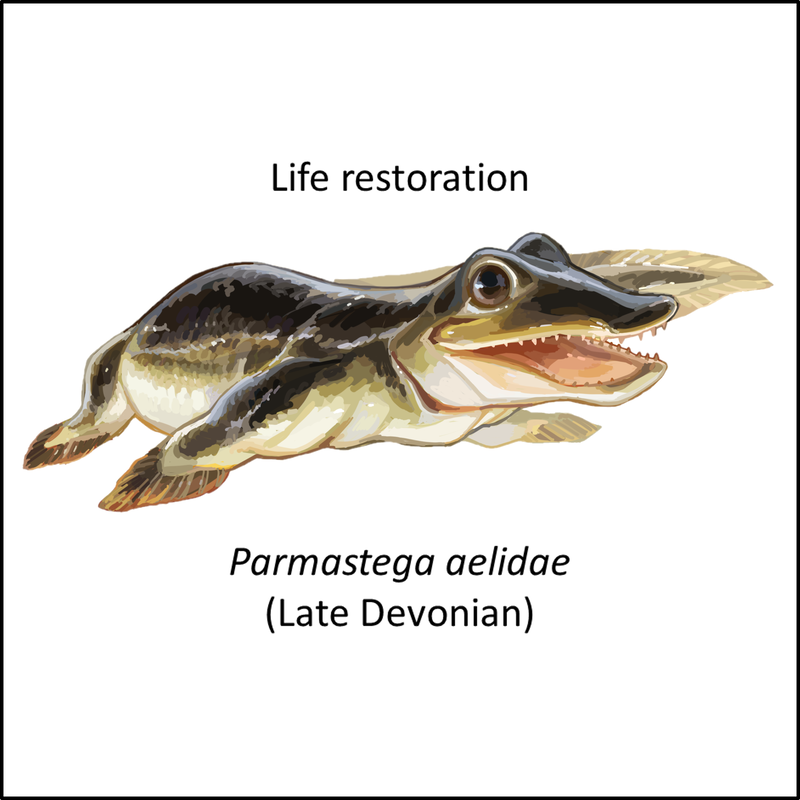















Names in red indicate that the fossil is younger than the oldest known crown-group fossil.
Figure 3. Images of stem-group tetrapods
This set of images illustrates the transition from fishes to tetrapods. The pictures are placed in order from most basal to most crownward in the stem group. Note the progressive change from fish-like (up to Eusthenodon) to increasingly lizard-like form (from Ventastega to Greererpeton) with a change from fins to limbs with digits (e.g. Acanthostega). However, this record is undoubtedly incomplete, because footprints interpreted to have been left by limbed tetrapods have been found in the Middle Devonian Valentia Slate Formation on Valentia Island in southwestern Ireland (Lucas, 2019); these are significantly older than the oldest known limbed tetrapods (e.g. Elginerpeton), which are of Late Devonian age (Ahlberg, 2018).
Some of the fossils illustrated above (with labels in red) are younger than the oldest crown group fossil; they represent descendants of undiscovered ancestors that would have separated from the stem line before the crown group appeared.
Much research is actively going on to understand the nature of the changes in the skeleton and musculature that allowed the stem-group tetrapods to move on land (see Ahlberg, 2018; Molnar et al, 2018, 2020 and 2021; Esteve-Altava et al, 2019; Ruta et al, 2018; Cloutier et al, 2020; Dickson et al, 2021; Hirasawa et al, 2022). The main developments are nicely summarized in the following figure from Ahlberg (2018):
Some of the fossils illustrated above (with labels in red) are younger than the oldest crown group fossil; they represent descendants of undiscovered ancestors that would have separated from the stem line before the crown group appeared.
Much research is actively going on to understand the nature of the changes in the skeleton and musculature that allowed the stem-group tetrapods to move on land (see Ahlberg, 2018; Molnar et al, 2018, 2020 and 2021; Esteve-Altava et al, 2019; Ruta et al, 2018; Cloutier et al, 2020; Dickson et al, 2021; Hirasawa et al, 2022). The main developments are nicely summarized in the following figure from Ahlberg (2018):
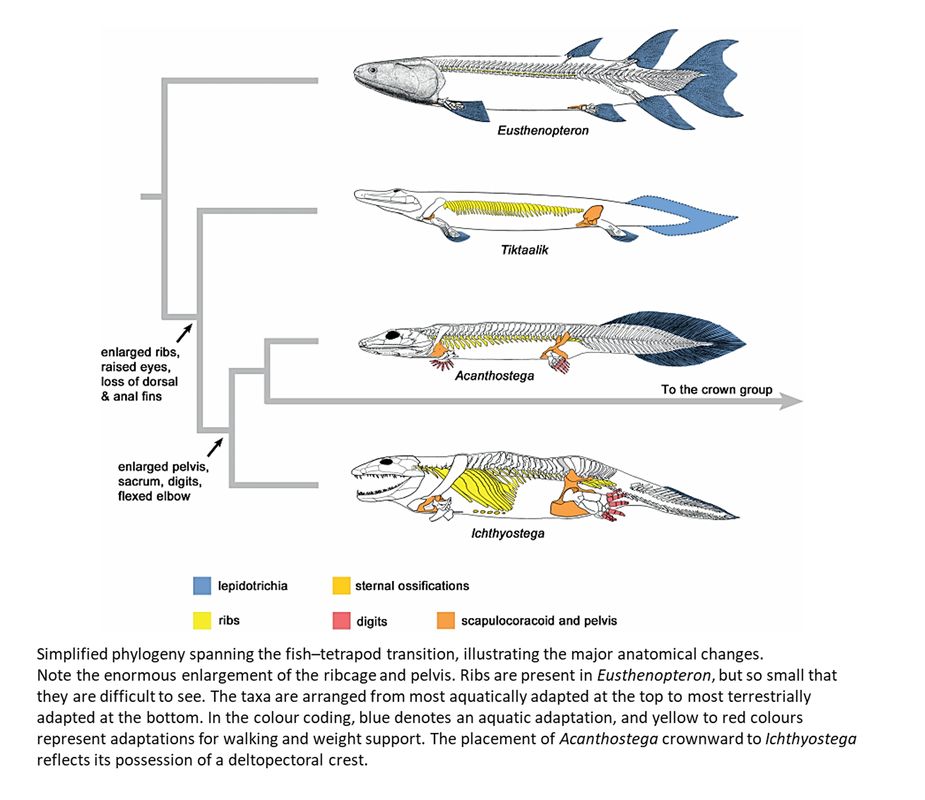
Figure 4. Taken from Open Access article by Ahlberg (2018)
One issue that remains unresolved is the relative timing of the appearance of limbs and the transition to a terrestrial lifestyle. Recent studies suggest that limbs developed first as a means of locomotion in or under water, and only later began to support weight on land (Lennie et al, 2021; Molnar et al, 2021):
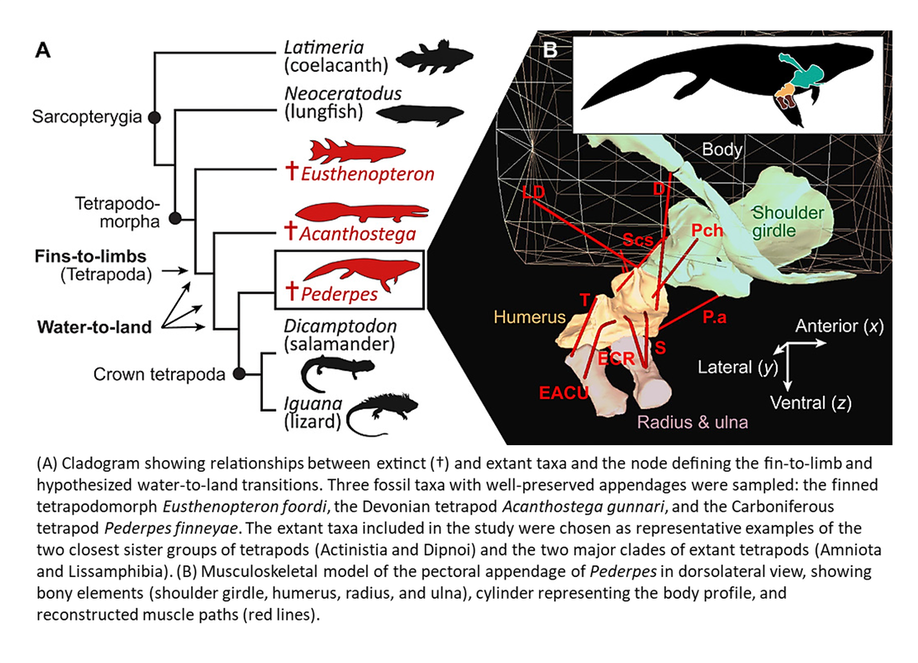
Figure 5. Taken from Open Access article by Molnar et al (2021)
This interpretation suggests that the earliest trackways do not necessarily imply locomotion on land; perhaps the trackways were made in an aquatic environment and subsequently exposed to air (Lennie et al, 2021).
Some idea of the nature of the transition from the stem group to the crown group of the tetrapods can be derived from a comparison of the above images with the examples of early crown-Tetrapoda shown below:
Some idea of the nature of the transition from the stem group to the crown group of the tetrapods can be derived from a comparison of the above images with the examples of early crown-Tetrapoda shown below:

Figure 6. Examples of early crown-Tetrapoda
The above time tree (Figure 3) indicates that the tetrapod stem group developed from Early Devonian to Early Carboniferous time, representing a stem-to-crown transition of at least 65 million years. However, the stem-group genera appeared at the greatest rate during the 24 million years occupied by the Late Devonian:
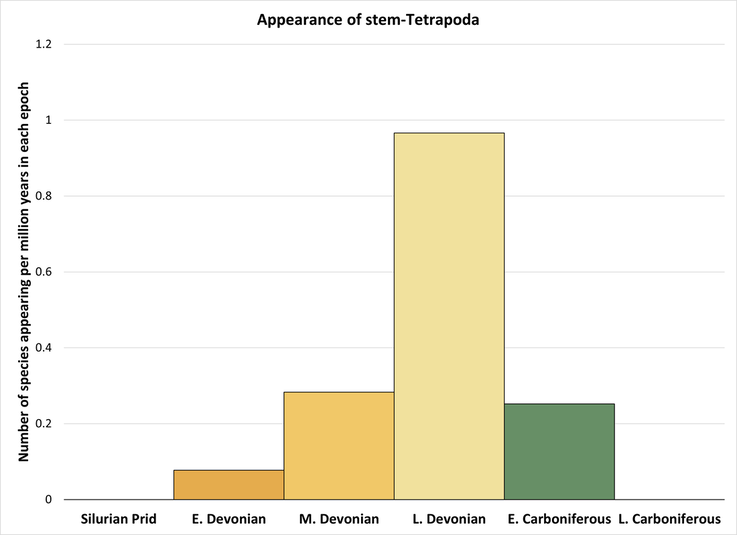
Figure 7. Rate of appearance of stem-group tetrapods (predating the crown group and including only the genera shown in Figure 3)
References
Ahlberg, P. E. (2018). Follow the footprints and mind the gaps: a new look at the origin of tetrapods. Earth and Environmental Science Transactions of the Royal Society of Edinburgh, 109(1-2), 115-137.
Ahlberg, P. E., & Clack, J. A. (2020). The smallest known Devonian tetrapod shows unexpectedly derived features. R. Soc. Open Sci.
7: 192117. http://dx.doi.org/10.1098/rsos.192117.
Benton, M. J. (2015). Vertebrate Palaeontology - Fourth edition. John Wiley & Sons, 468 pages.
Clack, J. A., Smithson, T. R., & Ruta, M. (2022). A Mississippian (early Carboniferous) tetrapod showing early diversification of the hindlimbs. Communications biology, 5(1), 1-10.
Cloutier, R., Clement, A. M., Lee, M. S., Noel, R., Bechard, I., Roy, V., & Long, J. A. (2020). Elpistostege and the origin of the vertebrate hand. Nature, 579(7800), 549-554.
Dickson, B. V., Clack, J. A., Smithson, T. R., & Pierce, S. E. (2021). Functional adaptive landscapes predict terrestrial capacity at the origin of limbs. Nature, 589(7841), 242-245.
Esteve-Altava, B., Pierce, S. E., Molnar, J. L., Johnston, P., Diogo, R., & Hutchinson, J. R. (2019). Evolutionary parallelisms of pectoral and pelvic network-anatomy from fins to limbs. Science advances, 5(5), eaau7459.
Hirasawa, T., Hu, Y., Uesugi, K., Hoshino, M., Manabe, M., & Kuratani, S. (2022). Morphology of Palaeospondylus shows affinity to tetrapod ancestors. Nature, 606(7912), 109-112.
Hohn-Schulte, B., Preuschoft, H., Witzel, U., & Distler-Hoffmann, C. (2013). Biomechanics and functional preconditions for terrestrial lifestyle in basal tetrapods, with special consideration of Tiktaalik roseae. Historical Biology, 25(2), 167-181.
Lennie, K. I., Manske, S. L., Mansky, C. F., & Anderson, J. S. (2021). Locomotory behaviour of early tetrapods from Blue Beach, Nova Scotia, revealed by novel microanatomical analysis. Royal Society open science, 8(5), 210281.
Lu, J., Zhu, M., Long, J. A., Zhao, W., Senden, T. J., Jia, L., & Qiao, T. (2012). The earliest known stem-tetrapod from the Lower Devonian of China. Nature Communications, 3, 1160.
Lucas, S. G. (2019). An ichnological perspective on some major events of Paleozoic tetrapod evolution. Bollettino della Società Paleontologica Italiana, 58(3), 224.
Molnar, J. L., Diogo, R., Hutchinson, J. R., & Pierce, S. E. (2018). Reconstructing pectoral appendicular muscle anatomy in fossil fish and tetrapods over the fins‐to‐limbs transition. Biological Reviews, 93(2), 1077-1107.
Molnar, J. L., Diogo, R., Hutchinson, J. R., & Pierce, S. E. (2020). Evolution of hindlimb muscle anatomy across the tetrapod water‐to‐land transition, including comparisons with forelimb anatomy. The Anatomical Record, 303(2), 218-234.
Molnar, J. L., Hutchinson, J. R., Diogo, R., Clack, J. A., & Pierce, S. E. (2021). Evolution of forelimb musculoskeletal function across the fish-to-tetrapod transition. Science Advances, 7(4), eabd7457.
Ruta, M., Krieger, J., Angieczyk, K. D., & Wiils, M. A. (2018). The evolution of the tetrapod humerus: Morphometrics, disparity, and evolutionary rates. Earth and Environmental Science Transactions of the Royal Society of Edinburgh, 109(1-2), 351.
Sanchez, S., Tafforeau, P., & Ahlberg, P. E. (2014). The humerus of Eusthenopteron: a puzzling organization presaging the establishment of tetrapod limb bone marrow. Proceedings of the Royal Society B: Biological Sciences, 281(1782), 20140299.
Shubin, N. H., Daeschler, E. B., & Jenkins, F. A. (2006). The pectoral fin of Tiktaalik roseae and the origin of the tetrapod limb. Nature, 440(7085), 764-771.
Sookias, R. B., Böhmer, C., & Clack, J. A. (2014). Redescription and phylogenetic analysis of the mandible of an enigmatic Pennsylvanian (Late Carboniferous) tetrapod from Nova Scotia, and the lability of Meckelian jaw ossification. PloS one, 9(10), e109717.
Ahlberg, P. E., & Clack, J. A. (2020). The smallest known Devonian tetrapod shows unexpectedly derived features. R. Soc. Open Sci.
7: 192117. http://dx.doi.org/10.1098/rsos.192117.
Benton, M. J. (2015). Vertebrate Palaeontology - Fourth edition. John Wiley & Sons, 468 pages.
Clack, J. A., Smithson, T. R., & Ruta, M. (2022). A Mississippian (early Carboniferous) tetrapod showing early diversification of the hindlimbs. Communications biology, 5(1), 1-10.
Cloutier, R., Clement, A. M., Lee, M. S., Noel, R., Bechard, I., Roy, V., & Long, J. A. (2020). Elpistostege and the origin of the vertebrate hand. Nature, 579(7800), 549-554.
Dickson, B. V., Clack, J. A., Smithson, T. R., & Pierce, S. E. (2021). Functional adaptive landscapes predict terrestrial capacity at the origin of limbs. Nature, 589(7841), 242-245.
Esteve-Altava, B., Pierce, S. E., Molnar, J. L., Johnston, P., Diogo, R., & Hutchinson, J. R. (2019). Evolutionary parallelisms of pectoral and pelvic network-anatomy from fins to limbs. Science advances, 5(5), eaau7459.
Hirasawa, T., Hu, Y., Uesugi, K., Hoshino, M., Manabe, M., & Kuratani, S. (2022). Morphology of Palaeospondylus shows affinity to tetrapod ancestors. Nature, 606(7912), 109-112.
Hohn-Schulte, B., Preuschoft, H., Witzel, U., & Distler-Hoffmann, C. (2013). Biomechanics and functional preconditions for terrestrial lifestyle in basal tetrapods, with special consideration of Tiktaalik roseae. Historical Biology, 25(2), 167-181.
Lennie, K. I., Manske, S. L., Mansky, C. F., & Anderson, J. S. (2021). Locomotory behaviour of early tetrapods from Blue Beach, Nova Scotia, revealed by novel microanatomical analysis. Royal Society open science, 8(5), 210281.
Lu, J., Zhu, M., Long, J. A., Zhao, W., Senden, T. J., Jia, L., & Qiao, T. (2012). The earliest known stem-tetrapod from the Lower Devonian of China. Nature Communications, 3, 1160.
Lucas, S. G. (2019). An ichnological perspective on some major events of Paleozoic tetrapod evolution. Bollettino della Società Paleontologica Italiana, 58(3), 224.
Molnar, J. L., Diogo, R., Hutchinson, J. R., & Pierce, S. E. (2018). Reconstructing pectoral appendicular muscle anatomy in fossil fish and tetrapods over the fins‐to‐limbs transition. Biological Reviews, 93(2), 1077-1107.
Molnar, J. L., Diogo, R., Hutchinson, J. R., & Pierce, S. E. (2020). Evolution of hindlimb muscle anatomy across the tetrapod water‐to‐land transition, including comparisons with forelimb anatomy. The Anatomical Record, 303(2), 218-234.
Molnar, J. L., Hutchinson, J. R., Diogo, R., Clack, J. A., & Pierce, S. E. (2021). Evolution of forelimb musculoskeletal function across the fish-to-tetrapod transition. Science Advances, 7(4), eabd7457.
Ruta, M., Krieger, J., Angieczyk, K. D., & Wiils, M. A. (2018). The evolution of the tetrapod humerus: Morphometrics, disparity, and evolutionary rates. Earth and Environmental Science Transactions of the Royal Society of Edinburgh, 109(1-2), 351.
Sanchez, S., Tafforeau, P., & Ahlberg, P. E. (2014). The humerus of Eusthenopteron: a puzzling organization presaging the establishment of tetrapod limb bone marrow. Proceedings of the Royal Society B: Biological Sciences, 281(1782), 20140299.
Shubin, N. H., Daeschler, E. B., & Jenkins, F. A. (2006). The pectoral fin of Tiktaalik roseae and the origin of the tetrapod limb. Nature, 440(7085), 764-771.
Sookias, R. B., Böhmer, C., & Clack, J. A. (2014). Redescription and phylogenetic analysis of the mandible of an enigmatic Pennsylvanian (Late Carboniferous) tetrapod from Nova Scotia, and the lability of Meckelian jaw ossification. PloS one, 9(10), e109717.
Image credits – Stem-Tetrapoda
- Figure 3 (Tungsenia paradoxa): Nobu Tamura under a Creative Commons 3.0 Unported (CC BY-NC-ND 3.0) license
- Figure 3 (Gooloogongia loomesi): ДиБгд, CC BY-SA 4.0 <https://creativecommons.org/licenses/by-sa/4.0>, via Wikimedia Commons
- Figure 3 (Sauripterus taylori): Ghedoghedo, CC BY-SA 3.0 <https://creativecommons.org/licenses/by-sa/3.0>, via Wikimedia Commons
- Figure 3 (Rhizodus hibberti): DiBgd, CC BY-SA 4.0 <https://creativecommons.org/licenses/by-sa/4.0>, via Wikimedia Commons
- Figure 3 (Gyroptychius agassizi): Ghedoghedo, CC BY-SA 3.0 <https://creativecommons.org/licenses/by-sa/3.0>, via Wikimedia Commons
- Figure 3 (Gogonasus andrewsae, fossils): From Open Access article Trinajstic, K., Briggs, D. E., & Long, J. A. (2022). The Gogo Formation Lagerstätte: a view of Australia's first great barrier reef. Journal of the Geological Society, 179(1).
- Figure 3 (Gogonasus andrewsae, life restoration): Nobu Tamura under a Creative Commons 3.0 Unported (CC BY-NC-ND 3.0) license
- Figure 3 (Megalichthys hibberti): Nobu Tamura under Creative Commons Attribution- ShareAlike (CC BY-SA) license
- Figure 3 (Ectosteorhachis nitidus): GhedoghedoGhedoghedo, Public domain, via Wikimedia Commons
- Figure 3 (Osteolepis macrolepidotus, fossil): Daderot, CC0, via Wikimedia Commons
- Figure 3 (Osteolepis macrolepidotus, life restoration): Nobu Tamura under a Creative Commons 3.0 Unported (CC BY-NC-ND 3.0) license
- Figure 3 (Panderichthys rhombolepis, fossil): Ghedoghedo, CC BY-SA 4.0 <https://creativecommons.org/licenses/by-sa/4.0>, via Wikimedia Commons
- Figure 3 (Panderichthys rhombolepis, life restoration): Nobu Tamura under a Creative Commons 3.0 Unported (CC BY-NC-ND 3.0) license
- Figure 3 (Eusthenopteron foordi, fossil): Ghedoghedo, CC BY-SA 3.0 <https://creativecommons.org/licenses/by-sa/3.0>, via Wikimedia Commons
- Figure 3 (Eusthenopteron foordi, life restoration): Nobu Tamura under a Creative Commons 3.0 Unported (CC BY-NC-ND 3.0) license
- Figure 3 (Elpistostege watsoni): Placoderm2, CC BY-SA 4.0 <https://creativecommons.org/licenses/by-sa/4.0>, via Wikimedia Commons
- Figure 3 (Tiktaalik roseae, fossil): Ghedoghedo, CC BY-SA 3.0 <https://creativecommons.org/licenses/by-sa/3.0>, via Wikimedia Commons
- Figure 3 (Tiktaalik roseae, life restoration): Nobu Tamura under Creative Commons Attribution- ShareAlike (CC BY-SA) license
- Figure 3 (Parmastega aelidae): Entelognathus, CC BY-SA 4.0 <https://creativecommons.org/licenses/by-sa/4.0>, via Wikimedia Commons
- Figure 3 (Ventastega curonica): Nobu Tamura under a Creative Commons 3.0 Unported (CC BY-NC-ND 3.0) license
- Figure 3 (Acanthostega gunnari, fossil): Ryan Somma, CC BY-SA 2.0 <https://creativecommons.org/licenses/by-sa/2.0>, via Wikimedia Commons
- Figure 3 (Acanthostega gunnari, life restoration): Nobu Tamura under a Creative Commons 3.0 Unported (CC BY-NC-ND 3.0) license
- Figure 3 (Elginerpeton pancheni): Nobu Tamura under a Creative Commons 3.0 Unported (CC BY-NC-ND 3.0) license
- Figure 3 (Ichthyostega sp.): OlegTarabanov, CC BY-SA 3.0 <https://creativecommons.org/licenses/by-sa/3.0>, via Wikimedia Commons
- Figure 3 (Ichthyostega stensioei): Nobu Tamura under a Creative Commons 3.0 Unported (CC BY-NC-ND 3.0) license
- Figure 3 (Ymeria denticulata): FunkMonk (Michael B. H.), CC BY-SA 3.0 <https://creativecommons.org/licenses/by-sa/3.0>, via Wikimedia Commons
- Figure 3 (Whatcheeria deltae): Nobu Tamura under Creative Commons Attribution- ShareAlike (CC BY-SA) license
- Figure 3 (Pederpes finneyae, fossil): From Open Access article Clack, J. A. (2009). The fish–tetrapod transition: new fossils and interpretations. Evolution: Education and Outreach, 2(2), 213-223.
- Figure 3 (Pederpes finneyae, life restoration): Nobu Tamura under Creative Commons Attribution- ShareAlike (CC BY-SA) license
- Figure 3 (Greererpeton burkemorani, fossil): Tim Evanson, CC BY-SA 2.0 <https://creativecommons.org/licenses/by-sa/2.0>, via Wikimedia Commons
- Figure 3 (Greererpeton burkemorani, life restoration): Nobu Tamura under a Creative Commons 3.0 Unported (CC BY-NC-ND 3.0) license
- Figure 3 (Crassigyrinus scoticus): Nobu Tamura under a Creative Commons 3.0 Unported (CC BY-NC-ND 3.0) license
- Figure 3 (Megalocephalus pachycephalus): Dmitry Bogdanov}}., CC BY 3.0 <https://creativecommons.org/licenses/by/3.0>, via Wikimedia Commons
- Figure 3 (Baphetes kirkbyi): British Geological Survey, licensed under a Creative Commons Attribution-NonCommercial-ShareAlike 3.0 Unported License.
- Figure 6 (Balanerpeton woodi): Nobu Tamura under a Creative Commons 3.0 Unported (CC BY-NC-ND 3.0) license
- Figure 6 (Dendrerpeton sp.): ДиБгд, Public domain, via Wikimedia Commons
- Figure 6 (Caerorhachis bairdi): Богданов [email protected], Public domain, via Wikimedia Commons
- Figure 6 (Silvanerpeton miripedes): Dmitry Bogdanov, CC BY-SA 3.0 <https://creativecommons.org/licenses/by-sa/3.0>, via Wikimedia Commons