This page deals with the stem group of the bichirs and reedfish (Class Cladistia (using the more widely-used name rather than "Cladistei" as proposed by Ruggiero et al, 2015), superclass Actinopterygii). These fishes, represented at the present time by about 14 species, all belong to one family, the Polypteridae, in the order Polypteriformes. They are elongated in form with fleshy lobes in the pectoral fins, rounded tails, thick ganoid (hard, rhombic, enameled) scales, and functional lungs for breathing air in addition to rudimentary gills (Encyclopedia Britannica).
The stem-Cladistia are rather sparsely represented in the fossil record and their phylogenetic relationships have proved difficult to determine, as exemplified in the phylogenetic time tree below, which shows all the stem-group species as well as the crown group deriving from a single node in a polytomy:
The stem-Cladistia are rather sparsely represented in the fossil record and their phylogenetic relationships have proved difficult to determine, as exemplified in the phylogenetic time tree below, which shows all the stem-group species as well as the crown group deriving from a single node in a polytomy:
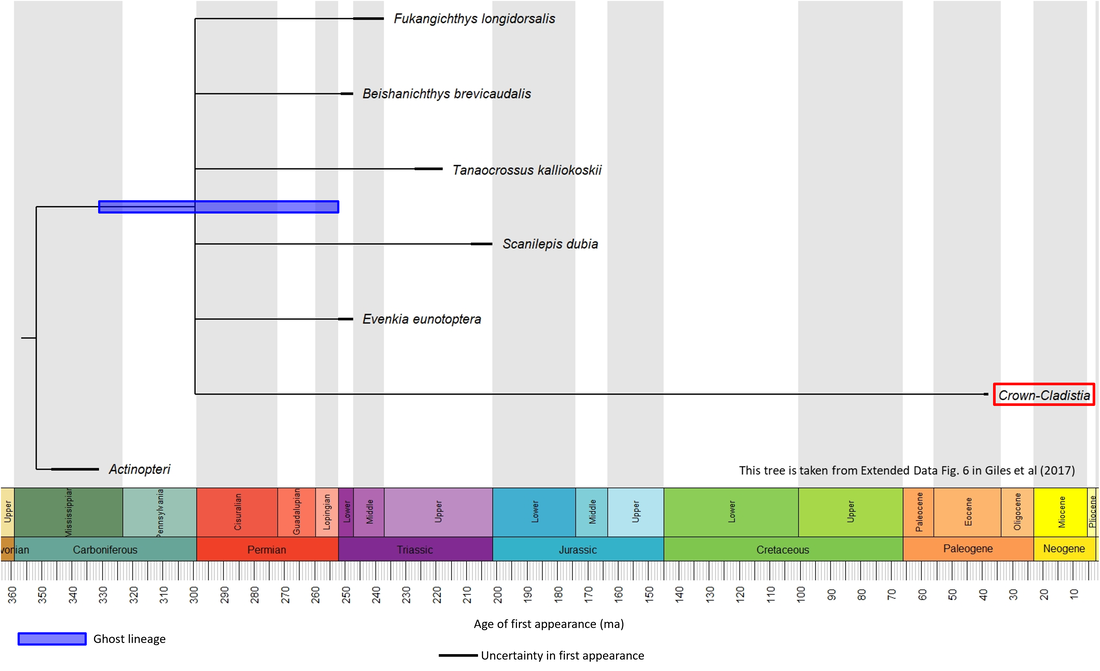
Figure 1. Time tree of the stem-Cladistia
The oldest known member of the stem-Cladistia is Evenkia eunotoptera, described from Early Triassic sediments in the Tunguska Coal basin, Yenisei, Siberia (Berg, 1941; Giles et al, 2017). No images are available in the public domain for this species or for any other stem-group cladistian except Tanaocrossus kalliokoskii, which is illustrated below:

Figure 2. Tanaocrossus kalliokoskii from the Late Triassic Chinle Formation of Utah
The oldest known member of the crown-Cladistia is Polypterus sp., found in the Middle Eocene New Idam Unit at Dur At-Talah, in the Sirt Basin, southeastern Libya (Otero et al, 2015). No public-domain image, apart from teeth, scales and fragments of jawbones, is available.
Making allowance for the long ghost lineage on the cladistian stem line (Figure 1), the time between the origin of the cladistian stem group and the initiation of the crown group was at least 290 million years, from Early Carboniferous to Eocene time. However, the long gap of at least 162 million years between the youngest known stem-group member (Scanilepis dubia) and the oldest crown group representative suggests the possibility that younger stem-group fossils, or older crown-Cladistia, remain to be found.
Making allowance for the long ghost lineage on the cladistian stem line (Figure 1), the time between the origin of the cladistian stem group and the initiation of the crown group was at least 290 million years, from Early Carboniferous to Eocene time. However, the long gap of at least 162 million years between the youngest known stem-group member (Scanilepis dubia) and the oldest crown group representative suggests the possibility that younger stem-group fossils, or older crown-Cladistia, remain to be found.
References
Berg, L. S. (1941). Lower Triassic fishes of the Tunguska coal basin, Yenisei, Siberia. Bulletin of the Academy of Sciences USSR, Biological Sciences, 3, 414-419.
Gardiner, B. G., Schaeffer, B., & Masserie, J. A. (2005). A review of the lower actinopterygian phylogeny. Zoological Journal of the Linnean Society, 144(4), 511-525.
Giles, S., Xu, G. H., Near, T. J., & Friedman, M. (2017). Early members of ‘living fossil’ lineage imply later origin of modern ray-finned fishes. Nature, 549(7671), 265.
Lu, J., Giles, S., Friedman, M., den Blaauwen, J. L., & Zhu, M. (2016). The oldest actinopterygian highlights the cryptic early history of the hyperdiverse ray-finned fishes. Current Biology, 26(12), 1602-1608.
Otero, O., Pinton, A., Cappetta, H., Adnet, S., Valentin, X., Salem, M., & Jaeger, J. J. (2015). A fish assemblage from the Middle Eocene from Libya (Dur At-Talah) and the earliest record of modern African fish genera. PLoS One, 10(12), e0144358.
Ruggiero, M. A., Gordon, D. P., Orrell, T. M., Bailly, N., Bourgoin, T., Brusca, R. C., ... & Kirk, P. M. (2015). A higher level classification of all living organisms. PloS one, 10(4), e0119248.
Stack, J., & Gottfried, M. D. (2022). A new, exceptionally well-preserved Permian actinopterygian fish from the Minnekahta Limestone of South Dakota, USA. Journal of Systematic Palaeontology, 1-32.
Gardiner, B. G., Schaeffer, B., & Masserie, J. A. (2005). A review of the lower actinopterygian phylogeny. Zoological Journal of the Linnean Society, 144(4), 511-525.
Giles, S., Xu, G. H., Near, T. J., & Friedman, M. (2017). Early members of ‘living fossil’ lineage imply later origin of modern ray-finned fishes. Nature, 549(7671), 265.
Lu, J., Giles, S., Friedman, M., den Blaauwen, J. L., & Zhu, M. (2016). The oldest actinopterygian highlights the cryptic early history of the hyperdiverse ray-finned fishes. Current Biology, 26(12), 1602-1608.
Otero, O., Pinton, A., Cappetta, H., Adnet, S., Valentin, X., Salem, M., & Jaeger, J. J. (2015). A fish assemblage from the Middle Eocene from Libya (Dur At-Talah) and the earliest record of modern African fish genera. PLoS One, 10(12), e0144358.
Ruggiero, M. A., Gordon, D. P., Orrell, T. M., Bailly, N., Bourgoin, T., Brusca, R. C., ... & Kirk, P. M. (2015). A higher level classification of all living organisms. PloS one, 10(4), e0119248.
Stack, J., & Gottfried, M. D. (2022). A new, exceptionally well-preserved Permian actinopterygian fish from the Minnekahta Limestone of South Dakota, USA. Journal of Systematic Palaeontology, 1-32.
Image credits - Stem-Cladistia
- Header (Polypterus delhezi): ばぶじ~, CC BY-SA 4.0 <https://creativecommons.org/licenses/by-sa/4.0>, via Wikimedia Commons
- Figure 2: Open Access article Hunt-Foster, R., Lockley, M., Milner, A., Foster, J., Matthews, N., Breithaupt, B., & Smith, J. (2016). Tracking dinosaurs in BLM canyon country, Utah. Geology of the intermountain west, 3, 67-100.