The odd-toed ungulates (order Perissodactyla, infraclass Eutheria) are a clade of animals characterized by the possession of either one or three hoofed toes on each hindfoot (Encyclopædia Britannica). The extant members of the crown group are divided into three families: the Equidae (horses and their relatives), Rhinocerotidae (rhinoceroses), and Tapiridae (tapirs).
Consensus has not yet been reached concerning the phylogeny of the stem-Perissodactyla. There are significant differences between the phylogenetic studies that have been published in recent years (e.g. Cooper et al, 2014; Rose et al, 2014; Chimento and Agnolin, 2020). The most recent one is used as the basis for the following time tree:
Consensus has not yet been reached concerning the phylogeny of the stem-Perissodactyla. There are significant differences between the phylogenetic studies that have been published in recent years (e.g. Cooper et al, 2014; Rose et al, 2014; Chimento and Agnolin, 2020). The most recent one is used as the basis for the following time tree:
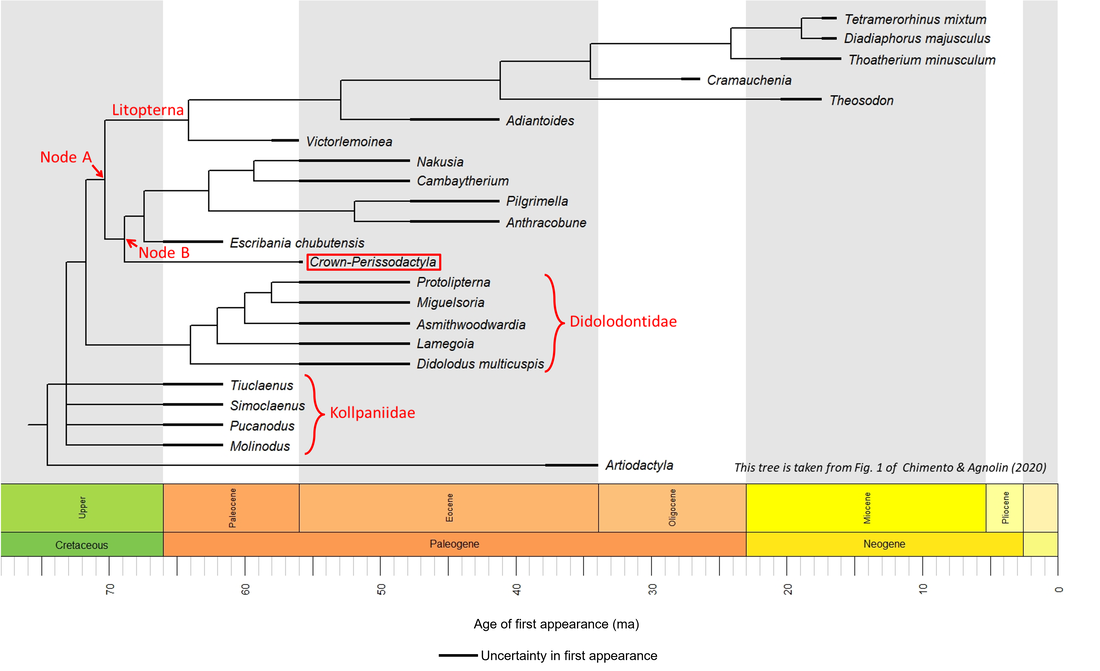
Figure 1. Phylogenetic time tree of the stem-Perissodactyla
As illustrated above, a number of fossils represent the oldest known members of the perissodactylan stem group: Escribania chubutensis, Molinodus, Pucanodus, Simoclaenus and Tiuclaenus, which are all of Danian (Early Paleocene) age. Escribania chubutensis is described from the Salamanca Formation at Punta Peligro in Chubut, Argentina (Bonaparte, 1993), while the other four genera are described from the Santa Lucía Formation of Bolivia. Escribania is illustrated below, together with other stem-group fossils for which images are available (for a larger view, click on image):
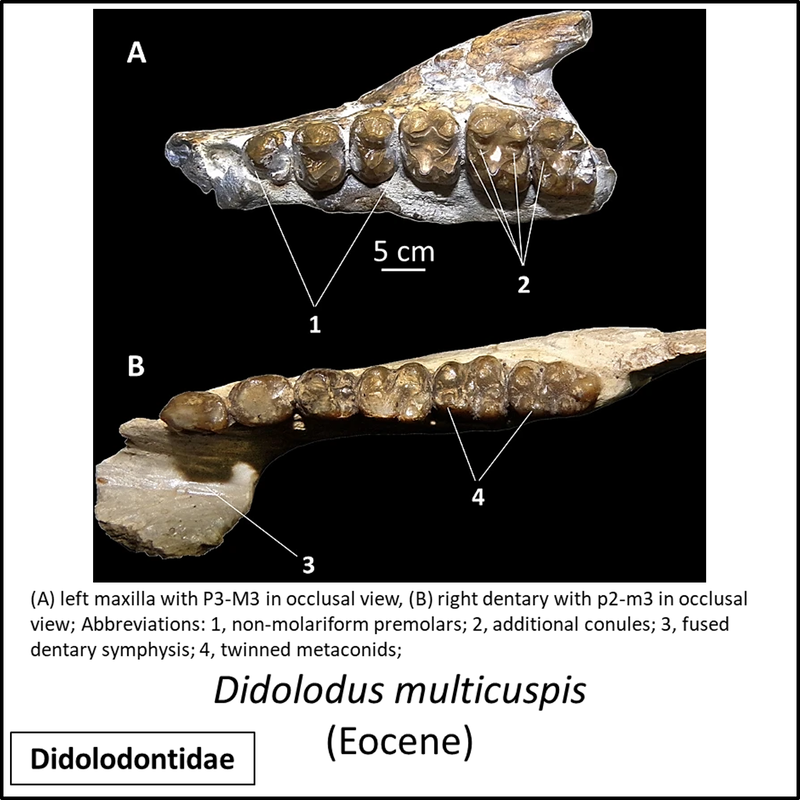













Names in red indicate that the fossil is younger than the oldest known crown-group fossil.
Figure 2. Images of stem-Perissodactyla
The above images are ordered from most basal to most crownward, but most of the images are of partial skeletal remains and no obvious trends in general appearance can be seen.
Note that several of the species shown above (represented by names in red) post-date the appearance of the crown group; these represent branches of the stem line that continued to evolve after the crown-group had appeared.
There is some controversy over the identity of the oldest member of the crown-Perissodactyla. It was suggested to be Hyracotherium sp. (Emerling et al, 2015; Foley et al, 2016), but that interpretation has been questioned by several researchers (e.g Bai et al, 2018; Upham et al, 2019). We will follow the suggestion of Westbury et al (2017) that the oldest known crown-group perissodactylan is the stem horse Sifrhippus sandrae. This species is described from several sites in the Early Eocene (North American Land Mammal stage Wasatchian Wa0) of Wyoming (Secord et al, 2012). A skeleton and a life restoration are shown below (for a larger view, click on image):
Note that several of the species shown above (represented by names in red) post-date the appearance of the crown group; these represent branches of the stem line that continued to evolve after the crown-group had appeared.
There is some controversy over the identity of the oldest member of the crown-Perissodactyla. It was suggested to be Hyracotherium sp. (Emerling et al, 2015; Foley et al, 2016), but that interpretation has been questioned by several researchers (e.g Bai et al, 2018; Upham et al, 2019). We will follow the suggestion of Westbury et al (2017) that the oldest known crown-group perissodactylan is the stem horse Sifrhippus sandrae. This species is described from several sites in the Early Eocene (North American Land Mammal stage Wasatchian Wa0) of Wyoming (Secord et al, 2012). A skeleton and a life restoration are shown below (for a larger view, click on image):


Figure 3. Sifrhippus, the oldest known crown-group odd-toed ungulate
The available fossil data indicate that the perissodactylan stem group developed from Paleocene to Early Eocene time, representing a minimum stem-to-crown transition period of only 5.6 million years.
References
Bai, B., Wang, Y. Q., & Meng, J. (2018). The divergence and dispersal of early perissodactyls as evidenced by early Eocene equids from Asia. Communications Biology, 1(1), 1-10.
Bonaparte, J. F. (1993). La fauna local de Punta Peligro, Paleoceno inferior, de la provincia del Chubut, Patagonia, Argentina (Vol. 14). Evolutionary Monographs.
Chimento, N. R., & Agnolin, F. L. (2020). Phylogenetic tree of Litopterna and Perissodactyla indicates a complex early history of hoofed mammals. Nature scientific reports, 10(1), 1-10.
Cooper, L. N., Seiffert, E. R., Clementz, M., Madar, S. I., Bajpai, S., Hussain, S. T., & Thewissen, J. G. (2014). Anthracobunids from the middle Eocene of India and Pakistan are stem perissodactyls. PloS one, 9(10), e109232.
Emerling, C. A., Huynh, H. T., Nguyen, M. A., Meredith, R. W., & Springer, M. S. (2015). Spectral shifts of mammalian ultraviolet-sensitive pigments (short wavelength-sensitive opsin 1) are associated with eye length and photic niche evolution. Proceedings of the Royal Society B: Biological Sciences, 282(1819), 20151817.
Foley, N. M., Springer, M. S., & Teeling, E. C. (2016). Mammal madness: is the mammal tree of life not yet resolved?. Philosophical Transactions of the Royal Society B: Biological Sciences, 371(1699), 20150140.
Rose, K. D., Holbrook, L. T., Rana, R. S., Kumar, K., Jones, K. E., Ahrens, H. E., ... & Smith, T. (2014). Early Eocene fossils suggest that the mammalian order Perissodactyla originated in India. Nature communications, 5(1), 1-9.
Secord, R., Bloch, J. I., Chester, S. G., Boyer, D. M., Wood, A. R., Wing, S. L., ... & Krigbaum, J. (2012). Evolution of the earliest horses driven by climate change in the Paleocene-Eocene Thermal Maximum. Science, 335(6071), 959-962.
Upham, N. S., Esselstyn, J. A., & Jetz, W. (2019). Inferring the mammal tree: species-level sets of phylogenies for questions in ecology, evolution, and conservation. PLoS biology, 17(12), e3000494.
Westbury, M., Baleka, S., Barlow, A., Hartmann, S., Paijmans, J. L., Kramarz, A., ... & Hofreiter, M. (2017). A mitogenomic timetree for Darwin’s enigmatic South American mammal Macrauchenia patachonica. Nature Communications, 8(1), 1-8.
Bonaparte, J. F. (1993). La fauna local de Punta Peligro, Paleoceno inferior, de la provincia del Chubut, Patagonia, Argentina (Vol. 14). Evolutionary Monographs.
Chimento, N. R., & Agnolin, F. L. (2020). Phylogenetic tree of Litopterna and Perissodactyla indicates a complex early history of hoofed mammals. Nature scientific reports, 10(1), 1-10.
Cooper, L. N., Seiffert, E. R., Clementz, M., Madar, S. I., Bajpai, S., Hussain, S. T., & Thewissen, J. G. (2014). Anthracobunids from the middle Eocene of India and Pakistan are stem perissodactyls. PloS one, 9(10), e109232.
Emerling, C. A., Huynh, H. T., Nguyen, M. A., Meredith, R. W., & Springer, M. S. (2015). Spectral shifts of mammalian ultraviolet-sensitive pigments (short wavelength-sensitive opsin 1) are associated with eye length and photic niche evolution. Proceedings of the Royal Society B: Biological Sciences, 282(1819), 20151817.
Foley, N. M., Springer, M. S., & Teeling, E. C. (2016). Mammal madness: is the mammal tree of life not yet resolved?. Philosophical Transactions of the Royal Society B: Biological Sciences, 371(1699), 20150140.
Rose, K. D., Holbrook, L. T., Rana, R. S., Kumar, K., Jones, K. E., Ahrens, H. E., ... & Smith, T. (2014). Early Eocene fossils suggest that the mammalian order Perissodactyla originated in India. Nature communications, 5(1), 1-9.
Secord, R., Bloch, J. I., Chester, S. G., Boyer, D. M., Wood, A. R., Wing, S. L., ... & Krigbaum, J. (2012). Evolution of the earliest horses driven by climate change in the Paleocene-Eocene Thermal Maximum. Science, 335(6071), 959-962.
Upham, N. S., Esselstyn, J. A., & Jetz, W. (2019). Inferring the mammal tree: species-level sets of phylogenies for questions in ecology, evolution, and conservation. PLoS biology, 17(12), e3000494.
Westbury, M., Baleka, S., Barlow, A., Hartmann, S., Paijmans, J. L., Kramarz, A., ... & Hofreiter, M. (2017). A mitogenomic timetree for Darwin’s enigmatic South American mammal Macrauchenia patachonica. Nature Communications, 8(1), 1-8.
Image credits – stem-Perissodactyla
- Header (black rhinos - Diceros bicornis - in Lewa, central Kenya): Harald Zimmer, CC BY-SA 3.0 <http://creativecommons.org/licenses/by-sa/3.0/>, via Wikimedia Commons
- Figure 2 (Didolodus multicuspis): Open Access article Chimento, N. R., & Agnolin, F. L. (2020). Phylogenetic tree of Litopterna and Perissodactyla indicates a complex early history of hoofed mammals. Nature Scientific reports, 10(1), 1-10.
- Figure 2 (Didolodus sp.): Pol.busquets, CC BY-SA 3.0 <https://creativecommons.org/licenses/by-sa/3.0>, via Wikimedia Commons
- Figure 2 (Adiantoides leali): Ghedo, CC BY-SA 4.0 <https://creativecommons.org/licenses/by-sa/4.0>, via Wikimedia Commons
- Figure 2 (Theosodon patagonicum): Ghedoghedo, CC BY-SA 3.0 <https://creativecommons.org/licenses/by-sa/3.0>, via Wikimedia Commons
- Figure 2 (Theosodon garretorum): Nobu Tamura, under Creative Commons Attribution- ShareAlike (CC BY-SA) license
- Figure 2 (Cramauchenia normalis): Smokeybjb, CC BY-SA 3.0 <https://creativecommons.org/licenses/by-sa/3.0>, via Wikimedia Commons
- Figure 2 (Thoatherium crepidatum): Ghedoghedo, CC BY-SA 3.0 <https://creativecommons.org/licenses/by-sa/3.0>, via Wikimedia Commons
- Figure 2 (Thoatherium minusculum): Charles R. Knight, Public domain, via Wikimedia Commons
- Figure 2 (Tetramerorhinus mixtum): Chimento, N. R., & Agnolin, F. L., CC BY 4.0 <https://creativecommons.org/licenses/by/4.0>, via Wikimedia Commons
- Figure 2 (Diadiaphorus robustus): Ghedoghedo, CC BY-SA 3.0 <https://creativecommons.org/licenses/by-sa/3.0>, via Wikimedia Commons
- Figure 2 (Diadiaphorus majusculus): Nobu Tamura, under Creative Commons Attribution- ShareAlike (CC BY-SA) license
- Figure 2 (Escribania chubutensis): Chimento, N. R., & Agnolin, F. L., CC BY 4.0 <https://creativecommons.org/licenses/by/4.0>, via Wikimedia Commons
- Figure 2 (Anthracobune spp.): Lisa Noelle Cooper, Erik R. Seiffert, Mark Clementz, Sandra I. Madar, Sunil Bajpai, S. Taseer Hussain, J. G. M. Thewissen, CC BY 4.0 <https://creativecommons.org/licenses/by/4.0>, via Wikimedia Commons
- Figure 2 (Anthracobune wardi): Nobu Tamura, under Creative Commons Attribution- ShareAlike (CC BY-SA) license
- Figure 3 (Sifrhippus sandrae): Jonathan Chen, CC BY-SA 4.0 <https://creativecommons.org/licenses/by-sa/4.0>, via Wikimedia Commons
- Figure 3 (Sifrhippus sp.): Eduard Solà, CC BY-SA 3.0 <https://creativecommons.org/licenses/by-sa/3.0>, via Wikimedia Commons