This page is the first in a series of pages that covers fossils associated with the stem group that leads to the crown group of the vertebrates (Subphylum Vertebrata, phylum Chordata) and with the stem groups of the clades within the crown group.
A summary of the phylogeny of the vertebrate clade is shown below:
A summary of the phylogeny of the vertebrate clade is shown below:
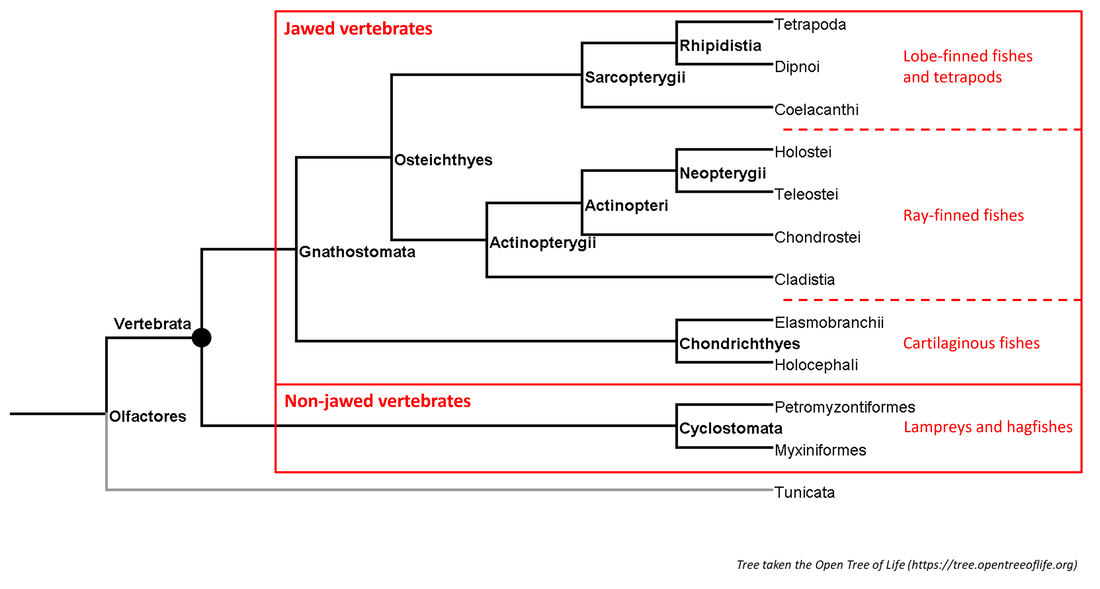
Figure 1. Summarized phylogenetic tree of the vertebrates
Each page of this website deals with a single branch of the phylogenetic tree. Links to the pages corresponding to the branches shown above (except for the Actinopteri and the Rhipidistia, for which no stem-group fossils are known) can be found under the Navigation tab above.
The remainder of this page deals with the stem-group vertebrates.
Few fossils have been interpreted as stem-Vertebrata. However, morphological analysis has resulted in three species being assigned to the vertebrate stem in a polytomy (Morris and Caron, 2014). Furthermore, in the phylogenetic tree presented by Miyashita et al (2019), the species are resolved in the stem-Vertebrata together with Haikouella lanceolata. However, given that the latter is now considered to be a yunnanozoan (Cong et al, 2015), it has been excluded from the phylogenetic time tree shown below:
The remainder of this page deals with the stem-group vertebrates.
Few fossils have been interpreted as stem-Vertebrata. However, morphological analysis has resulted in three species being assigned to the vertebrate stem in a polytomy (Morris and Caron, 2014). Furthermore, in the phylogenetic tree presented by Miyashita et al (2019), the species are resolved in the stem-Vertebrata together with Haikouella lanceolata. However, given that the latter is now considered to be a yunnanozoan (Cong et al, 2015), it has been excluded from the phylogenetic time tree shown below:
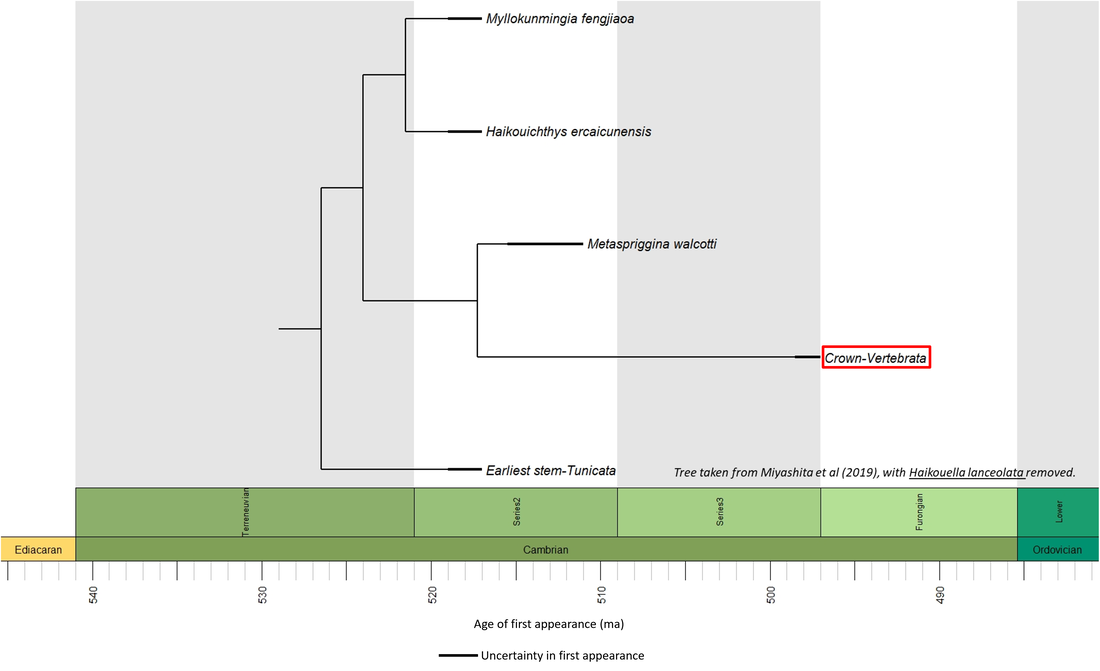
Figure 2. Phylogenetic time tree of the stem-Vertebrata
Images available in the public domain for the species represented in the above figure are shown below (click on image for a larger version):




Figure 3. Images of stem-group vertebrates
The above species are generally similar in being soft-bodied, with a notochord, prominent eyes and gill slits (Morris and Caron, 2014). They are the only known transitional fossils between the division of the vertebrate and tunicate stem lines and the appearance of the vertebrate crown group. Some idea of the nature of this transition can be gained by comparing the above images with the examples of early crown vertebrates shown below:
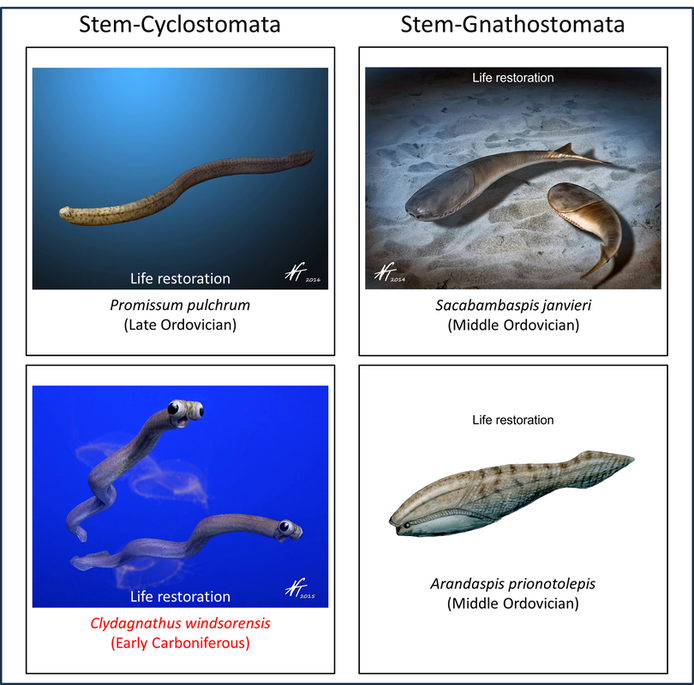
Names in red indicate that the fossil is younger than the oldest known crown-group fossil.
Figure 4. Examples of early crown-Vertebrata
This transition took place in the Early and Middle Cambrian over a period of about 20 million years, as indicated in the phylogenetic time tree above (Figure 2).
References
Benton, M. J. (2015). Vertebrate Palaeontology - Fourth edition. John Wiley & Sons, 468 pages.
Cong, P. Y., Hou, X. G., Aldridge, R. J., Purnell, M. A., & Li, Y. Z. (2015). New data on the palaeobiology of the enigmatic yunnanozoans from the Chengjiang Biota, Lower Cambrian, China. Palaeontology, 58(1), 45-70.
Miyashita, T., Coates, M. I., Farrar, R., Larson, P., Manning, P. L., Wogelius, R. A., ... & Currie, P. J. (2019). Hagfish from the Cretaceous Tethys Sea and a reconciliation of the morphological–molecular conflict in early vertebrate phylogeny. Proceedings of the National Academy of Sciences, 116(6), 2146-2151.
Morris, S. C., & Caron, J. B. (2014). A primitive fish from the Cambrian of North America. Nature, 512(7515), 419-422.
Cong, P. Y., Hou, X. G., Aldridge, R. J., Purnell, M. A., & Li, Y. Z. (2015). New data on the palaeobiology of the enigmatic yunnanozoans from the Chengjiang Biota, Lower Cambrian, China. Palaeontology, 58(1), 45-70.
Miyashita, T., Coates, M. I., Farrar, R., Larson, P., Manning, P. L., Wogelius, R. A., ... & Currie, P. J. (2019). Hagfish from the Cretaceous Tethys Sea and a reconciliation of the morphological–molecular conflict in early vertebrate phylogeny. Proceedings of the National Academy of Sciences, 116(6), 2146-2151.
Morris, S. C., & Caron, J. B. (2014). A primitive fish from the Cambrian of North America. Nature, 512(7515), 419-422.
Image credits - Stem-Vertebrata
- Header (Kentrosaurus aethiopicus skeleton): H. Zell [GFDL (http://www.gnu.org/copyleft/fdl.html) or CC BY-SA 3.0 (https://creativecommons.org/licenses/by-sa/3.0)], from Wikimedia CommonsFigure 2 (fossil): Degan Shu, Northwest University, Xi'an, China. Cropped by User:Andrew Dalby, Attribution, via Wikimedia Commons
- Figure 3 (fossil, Myllokunmingia fengjiaoa): Degan Shu, Northwest University, Xi'an, China. Cropped by User:Andrew Dalby, Attribution, via Wikimedia Commons
- Figure 3 (life restoration, Myllokunmingia fengjiaoa): Nobu Tamura under Creative Commons Attribution- ShareAlike (CC BY-SA) license
- Figure 3 (Haikouichthys ercaicunensis): Nobu Tamura under Creative Commons Attribution- ShareAlike (CC BY-SA) license
- Figure 3 (Metaspriggina walcotti): Nobu Tamura under Creative Commons Attribution- ShareAlike (CC BY-SA) license
- Figure 4 (Promissum pulchrum): Nobu Tamura under Creative Commons Attribution- ShareAlike (CC BY-SA) license
- Figure 4 (Clydagnathus windsorensis): Nobu Tamura under Creative Commons Attribution- ShareAlike (CC BY-SA) license
- Figure 4 (Sacabambaspis janvieri): Nobu Tamura, licensed under Creative Commons Attribution- ShareAlike (CC BY-SA)
- Figure 4 (Arandaspis prionotolepis): Nobu Tamura (http://spinops.blogspot.com) [CC BY-SA 3.0 (https://creativecommons.org/licenses/by-sa/3.0) or GFDL (http://www.gnu.org/copyleft/fdl.html)], from Wikimedia Commons